
广西不同气候条件下马尾松中龄林木本植物叶性状
2020-11-03 06:38潘小梅马姜明杨章旗卢忠耐零天旺
广西林业科学 2020年3期
菅 瑞,潘小梅,马姜明,杨章旗,郭 飞,卢忠耐,零天旺
(1.广西师范大学可持续发展创新研究院,广西桂林 541001;2.珍稀濒危动植物生态与环境保护教育部重点实验室,广西桂林 541001;3.广西壮族自治区林业科学研究院,广西南宁 530002;4.广西国有派阳山林场,广西宁明 532500;5.环江毛南族自治县华山林场,广西环江 547100;6.横县镇龙林场,广西横县 530327)
植物功能性状是由Díaz 和Cabido 在总结前人对植物性状研究的基础上首次提出的,是指一切对植物的定居、生存和适应有着潜在重要影响的,或与资源获取、利用和分配等相关的属性[1],这些属性是反映跨区域和跨尺度比较植物特征的指标,在植物与环境条件关系中发挥着重要作用[2],能单独或联合反映植物资源利用策略[3]。叶性状一直是生态学家研究的热点之一,近年来关于环境梯度的叶性状研究主要涉及纬度梯度[4]、降水梯度[5]和海拔梯度[6]等环境因子,从不同气候带的角度研究的较少。
马尾松(Pinus massoniana)广泛分布于我国17个省(自治区、直辖市)[7],因具有适应能力强、材质好、生长快、耐干旱、耐瘠薄和抗虫性强等特点,成为我国南方主要造林灭荒树种之一[8]。随着马尾松种植的规模化和集约化,马尾松得以速生,但同时地力衰退现象已成为马尾松可持续经营的主要障碍。本研究以不同气候条件下马尾松中龄林中的木本植物为研究对象,从叶性状特征入手,探究马尾松中龄林叶性状随环境梯度的变化规律及反映马尾松中龄林木本植物的关键叶性状,揭示马尾松中龄林木本植物沿环境梯度的生态策略,为预测环境变化对植物生存以及生态系统结构和功能的影响提供依据。
1 材料与方法
1.1 试验地概况
试验地位于广西宁明县国有派阳山林场(106°30'~107°15'E,21°15'~22°30'N),海拔200 ~800 m;属北热带季风气候,年均气温21.8 ℃,年均降水量1 475 mm;土壤以赤红壤为主。广西横县镇龙林场(109°08' ~109°19'E,23°02' ~23°08'N),海拔400 ~700 m;属南亚热带季风气候,年均气温21.5 ℃,年均降水量1 477.8 mm;土壤多为赤红壤,呈酸性或微酸性。广西环江毛南族自治县华山林场(108°06' ~108°38'E,25°6' ~25°31'N),海拔300 ~600 m;属中亚热带季风气候,年均气温19.8 ℃,年均降水量1 402 mm;土壤多为黄壤和红壤。
1.2 样地设置
在3 个试验地中分别选取1 个典型马尾松中龄林作为样地(表1)。选择无干扰、立地条件基本一致的地段设置样方,每个样地设置3个样方,样方面积为20 m×20 m,每个样方内设置4 个10 m×10 m的小样方。对各样方内的木本植物种类、树高、胸径和冠幅等进行群落学调查,并记录经纬度、海拔、坡向和坡度等情况。
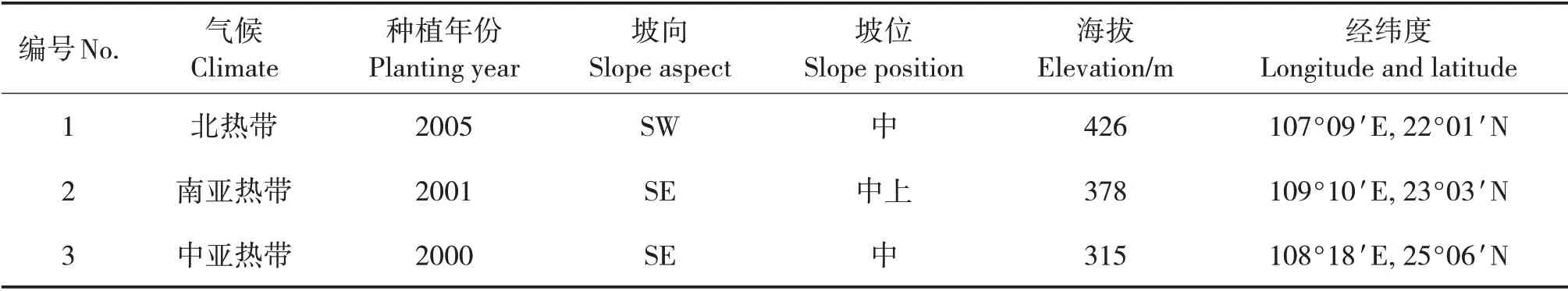
表1 样地基本情况Tab.1 Basic conditions of sample plots
1.3 试验方法
选取3 株生理形态健康成熟的植株,从不同方向采集10 片健康完整、大小相似的叶片,共30 片。去叶柄后置于湿润的滤纸中,装入自封袋后,放入有冰袋的保温包中储存。采集当天带回实验室后用水湿润,放在5 ℃的冰箱中储藏12 h,取出后迅速用吸水纸吸干叶片表面的水分,用电子天平称重(精确至0.000 1 g)并编号,得到叶饱和鲜重(SFW)。用游标卡尺测定叶厚度(LT,精确至0.1 mm),用叶面积仪测量叶面积(LA,cm2)。叶片样品置于60 ℃的烘箱烘至恒重后用电子天平测定叶干重(DW,g)。计算得到比叶面积(SLA,cm2/g)、叶干物质含量(LDMC,g/g)和叶组织密度(LTD,g/cm3),计算公式如下:

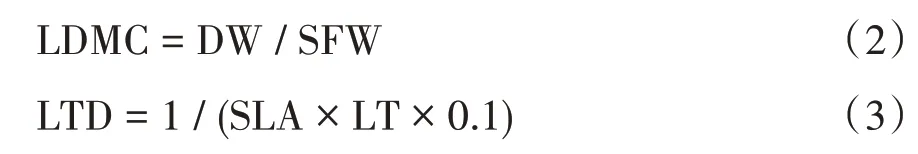
1.4 数据处理
对不同气候条件下马尾松中龄林乔木层和灌木层木本植物的6 个叶性状(LT、DW、LA、LDMC、SLA和LTD)进行Pearson相关性分析。
2 结果与分析
2.1 不同气候条件下马尾松中龄林木本植物叶性状的变化
LT、LDMC、DW 和LTD 随纬度的升高呈上升趋势;LA 和SLA 随纬度的升高呈下降趋势(图1)。
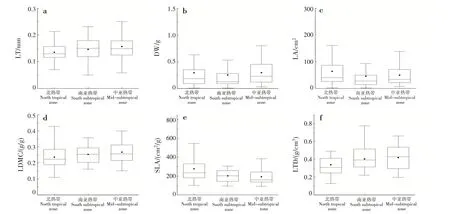
图1 叶性状在不同气候条件下的变化Fig.1 Changes of leaf traits in different climatic conditions
为进一步探讨叶片功能性状的变化规律,对乔木层和灌木层植物叶性状随纬度的变化趋势进行分析(图2)。乔木层LT、LDMC 和LTD 随纬度的升高呈上升趋势,DW、LA 和SLA 呈下降趋势;灌木层LT、LDMC 和LTD 随纬度的升高呈上升趋势,SLA 呈下降趋势,DW 和LA 变化趋势不明显。乔木层植物叶片LDMC和LTD在南亚热带和中亚热带均大于灌木层,SLA和LT则小于灌木层。
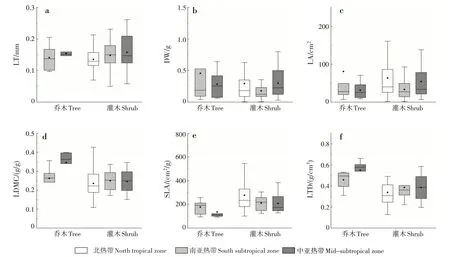
图2 不同气候条件下乔木层和灌木层植物叶性状的变化Fig.2 Changes of leaf traits in tree layer and shrub layer under different climatic conditions
2.2 相关性分析
北热带灌木层DW与LDMC、LA、LT和LTD,LDMC 与LT和LTD 以及LA 与LT均呈显著正相关;DW与SLA、LDMC 与LA 和SLA、LA 与LTD、LT与SLA 和LTD以及SLA与LTD均呈显著负相关(表2)。
南亚热带乔木层DW 与LA 和LT、LDMC 与LT和LTD、LA与LT和SLA均呈显著正相关;DW与LDMC 和LTD、LDMC 与LA 和SLA、LA 与LTD、LT 与SLA、SLA 与LTD 均呈显著负相关(表3)。灌木层DW 与LA 和LT、LDMC 与LTD、LA 与LT 均呈显著正相关;DW 与SLA,LDMC与LA、LT和SLA,LA与SLA和LTD,LT 与SLA 和LTD 以及SLA 与LTD 均呈显著负相关。
中亚热带乔木层DW 与LDMC、LA、LT 和LTD,LDMC 与LA、LT 和LTD 以及LA 与LT 和LTD 均呈显著正相关;SLA 与DW、LDMC、LA 和LT 均呈显著负相关(表4)。灌木层DW 与LDMC、LA 和LT,LDMC与LTD以及LA与LT和SLA均呈显著正相关;DW与SLA 和LTD、LDMC 与LA 和SLA、LA 与LTD、LT 与SLA和LTD以及SLA与LTD均呈显著负相关。
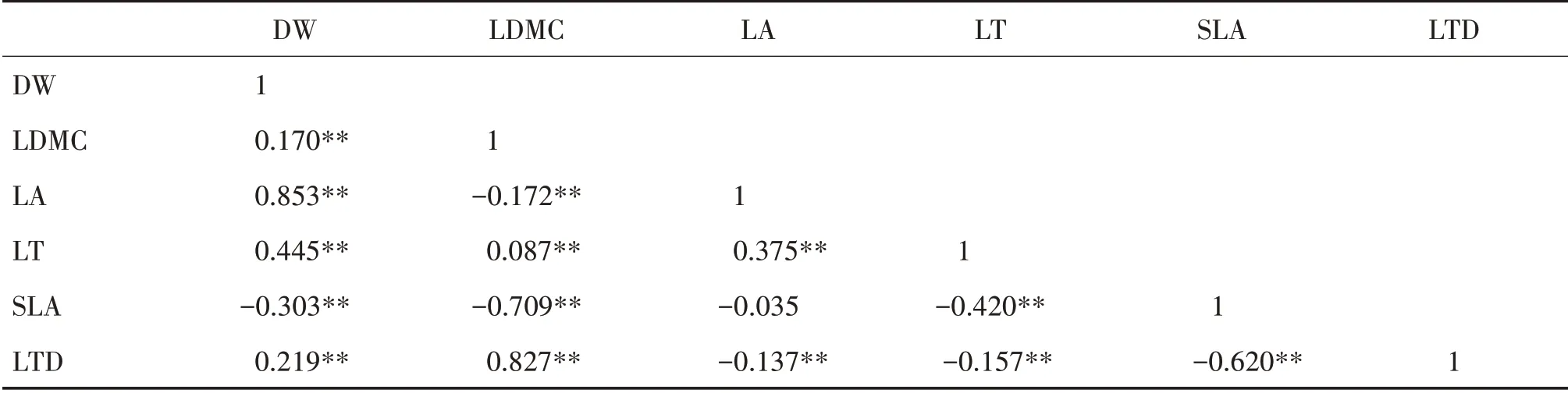
表2 北热带灌木层叶性状的Pearson相关性分析Tab.2 Pearson correlation analysis of leaf traits of shrub layer in north tropical zone
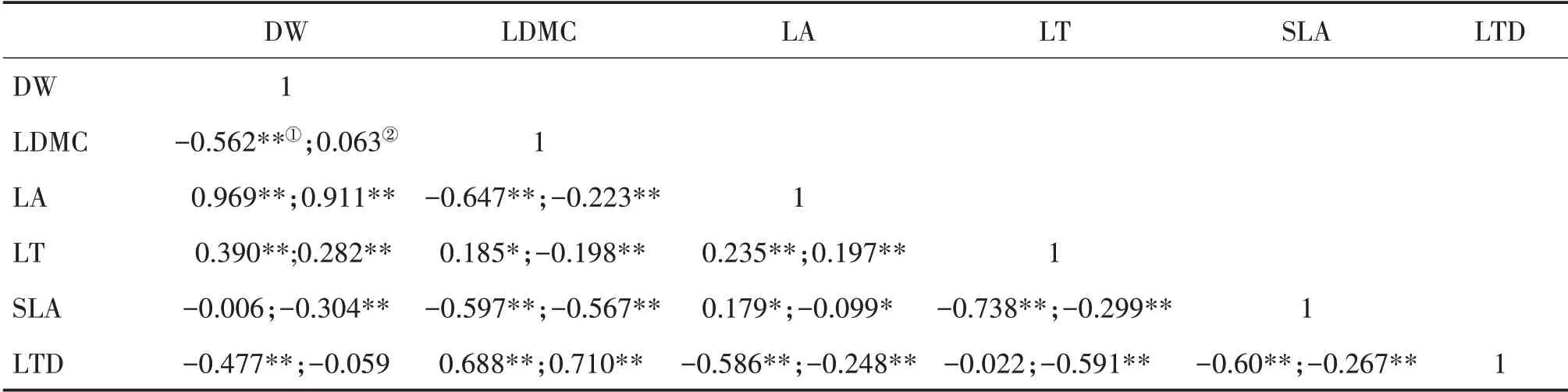
表3 南亚热带乔木层①和灌木层②叶性状的Pearson相关性分析Tab.3 Pearson correlation analysis of leaf traits of tree layer①and shrub layer②in south subtropical zone
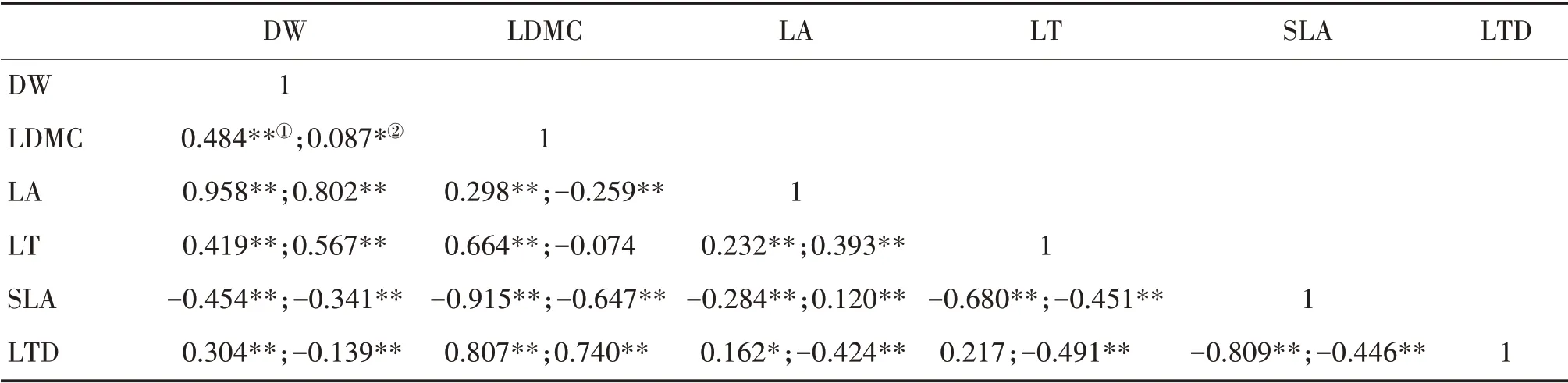
表4 中亚热带乔木层①和灌木层②叶性状的Pearson相关性分析Tab.4 Pearson correlation analysis of leaf traits of tree layer①and shrub layer②in mid-subtropical zone
2.3 主成分分析
南亚热带乔木层叶性状前2个主成分可以解释叶性状总方差的95%,其中PC1 可以解释总方差的60%,PC2可以解释总方差的35%;LA、LDMC和LTD对PC1 的贡献率较大;SLA 和LT 对PC2 的贡献率较大(图3a)。第一主成分突出反映植物对光的吸收和对碳的获取能力,第二主成分突出反映植物对光照和水分等异质性环境因子的资源利用策略。中亚热带乔木层叶性状前2个主成分可以解释叶性状总方差的90%,其中PC1 可以解释总方差的65%,PC2可以解释总方差的25%;SLA 和LDMC 对PC1的贡献率较大;LA对PC2的贡献率较大(图3b)。第一主成分突出反映植物抵御外界干扰的能力,第二主成分突出反映植物对光的吸收和对碳的获取能力。
北热带灌木层叶性状前2 个主成分可以解释叶性状总方差的82%,其中PC1 可以解释总方差的46%,PC2 可以解释总方差的36%;LDMC、LTD 和SLA 对PC1 的贡献率较大;LA 对PC2 的贡献率较大(图3c)。第一主成分突出反映植物抵御外界干扰的能力,第二主成分突出反映植物对光的吸收和对碳的获取能力。南亚热带灌木层叶性状前2个主成分可以解释叶性状总方差的77%,其中PC1 可以解释总方差的42%,PC2 可以解释总方差的35%;LA和LTD 对PC1 的贡献率较大;SLA 和LDMC 对PC2的贡献率较大(图3d)。第一主成分突出反映植物对光的吸收和对碳的获取能力,第二主成分突出反映植物抵御外界干扰的能力。中亚热带灌木层叶性状前2 个主成分可以解释叶性状总方差的83%,其中PC1 可以解释总方差的49%,PC2 可以解释总方差的34%;LA 和LTD 对PC1 的贡献率较大;SLA和LDMC 对PC2的贡献率较大(图3e)。第一主成分突出反映植物对光的吸收和对碳的获取能力,第二主成分突出反映植物抵御外界干扰的能力。
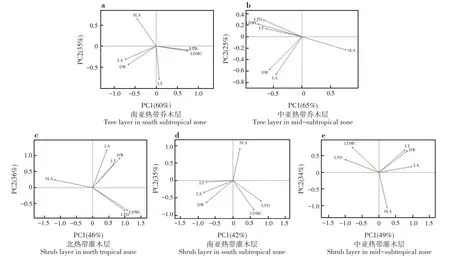
图3 叶性状的主成分分析Fig.3 Principal component analysis of leaf traits
3 结论与讨论
3.1 不同气候条件下植物叶性状的比较
LA随着纬度的升高,总体呈下降趋势。叶面积随着年均温的降低而减小[9],与本研究结果一致。纬度越高,气温越低,叶接受光能和同化二氧化碳的时间缩短,引起叶片小型化。小叶的呼吸和蒸腾成本更低[10],可以降低植株的维持消耗。LT 随着纬度的升高,呈上升趋势。植物叶片LT的增加是植物对低温适应的结果[11]。LDMC 能反映植物获取资源的能力,LDMC 越高,植物获取资源的能力越强[12]。LDMC 随着纬度的升高,呈上升趋势,SLA 则相反。吴漫玲等[13]对星斗山混交林木本植物不同海拔梯度的研究表明,随海拔升高,温度降低,LT 和LDMC 升高,LA 和SLA 降低,与本研究结果相似。低温是植物SLA 沿海拔梯度上升而下降的根本原因,在低温环境下,叶肉细胞增大和细胞壁特别是表皮细胞壁增厚,所以LT增加,SLA下降[14]。LTD随着纬度的升高,呈上升趋势。铁杆蒿(Tripolium pannonicum)在森林带(温度较高的植被带)拥有最低的LTD[15],与本研究结果一致。在高温环境下,通过增加LTD 可防止高温伤害[15]。
叶结构性状在不同生活型适应策略不同。乔木层和灌木层的LT和LTD随纬度的升高,均呈上升趋势,SLA 则相反。SLA 表征植物潜在相对生长速率或最大光合速率,能指示植物在不同环境下利用资源的能力[16]。本研究表明,乔木层SLA 均小于相应气候带的灌木层,这与莫燕华等[17]的研究结果一致。SLA 大的植物捕获光的能力强,通常光线越弱的冠层形成的叶子SLA 越大[18]。灌木层光线相对乔木层弱,SLA 的投资大。乔木层植物相对于灌木层植物具备更好地适应资源贫瘠的能力,灌木层植物则在获取资源(如光照)的能力方面相对较强[19]。本研究表明,乔木层LT 均小于对应的灌木层,乔木层LDMC 大于灌木层。黄小等[20]对不同生活型叶性状的研究表明,LT表现为乔木<灌木<木质藤本,LDMC 表现为乔木>灌木>木质藤本,与本研究结果一致。乔木层植物在叶片受到强光辐射时,通常会加大对LDMC的投资[17]。
3.2 植物叶性状之间的相关性
植物功能性状间紧密的相关关系普遍存在,这是由于物种间在生长发育及形成适应性机制的过程中相互作用,表现出相对应的权衡策略[21]。LA与LT在马尾松乔木层和灌木层均呈极显著正相关,这与滇中喀斯特植物[22]的研究结果一致。DW 与LA 和LT、LDMC 与LTD 在马尾松乔木层和灌木层均呈极显著正相关,这与滇中喀斯特植物[22]、琅岐岛优势植物[23]等研究结果一致,表明DW随LA和LT的增大而增大,LDMC 随LTD 的增大而增大,进一步表明LDMC 越大LTD 越高,叶也越坚韧,越能抵抗不良的生长物理因子[24]。
DW 与SLA、LDMC 与LA、LA 与LTD、LT 与SLA和LTD、SLA 与LTD 在马尾松灌木层均呈显著负相关,这与不同林龄马尾松人工林针叶植物[25]、井冈山鹿角杜鹃(Rhododendron latoucheae)群落植物[21]、桂林岩溶石山檵木(Loropetalum chinense)群落常见植物[17]和星斗山常绿落叶阔叶混交林木本植物[13]研究结果一致。SLA和LDMC是叶片最为关键的性状,能综合反映植物利用资源的能力[26],是植物适应生境的关键因子,具有重要的生态学意义。LDMC与SLA在马尾松乔木层和灌木层均表现为显著负相关,即随着LDMC 的增加,SLA 减小,使得叶片内部水分向叶片表面扩散的阻力增大,降低植物内部水分散失[27]。
3.3 植物叶性状适应策略
主成分分析表明,在6 个叶性状指标中,LA、LDMC、LTD 和LT 可作为反映南亚热带乔木层叶性状的重要指标,主要表征植物抵御外界不利环境和对水热环境条件适应的能力。LDMC 和LA 可作为反映中亚热带乔木层叶性状的重要指标,主要表征植物抵御外界干扰及不利环境的能力。乔木层植物叶片LDMC 均大于相应气候带的灌木层,表明乔木层植物具有“缓慢投资-收益”,即“保守型”叶经济谱的特点[28-29]。SLA、LTD 和LA 可作为反映北热带、南亚热带和中亚热带的马尾松中龄林灌木层叶性状的重要指标,表征植物获取资源及对干湿环境的适应能力。灌木层植物叶片SLA均大于相应气候带的乔木层,表明灌木层植物具有“快速投资-收益”,即“获取型”叶经济谱的特点[28-29]。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
锦绣·下旬刊(2020年12期)2020-01-03
绿色科技(2019年5期)2019-11-29
亚热带农业研究(2019年2期)2019-09-05
农村经济与科技(2018年15期)2018-05-14
森林工程(2018年5期)2018-05-14
中国粮油学报(2018年12期)2018-03-19
星星·诗歌原创(2017年5期)2017-08-13
中国果菜(2015年2期)2015-03-11
亚热带资源与环境学报(2015年4期)2015-01-22
