
高原红细胞增多症模型大鼠外周血EGLN1基因甲基化分析*
2020-11-03 07:59朱明明张培莉冀林华
中国高原医学与生物学杂志 2020年3期
杨 敏,朱明明,3,宋 康,4,张培莉,5,马 轶,廖 英,崔 森,3&,冀林华,5△
(1.青海大学医学院高原医学研究中心,青海 西宁 810001; 2.青海省高原医学应用基础重点实验室,青海 西宁 810001; 3.青海大学附属医院,青海 西宁 810001;4.青海省人民医院,青海 西宁 810001; 5.广州市花都区人民医院,广东 广州 510800)
目前的研究认为高原红细胞增多症(HAPC)病理改变的主要机制是对高原环境失适应致细胞内低氧使低氧诱导因子(hypoxia inducible factor,HIF)表达增加[1],围绕HIF上下游调节因子深入研究依然是HAPC的研究热点。研究发现,表观遗传学机制与人类高原低氧适应相关,是衔接环境与基因组改变的核心[2],提示我们可以以表观遗传学为新的视角去探讨HAPC的发病机制。
EGLN1位于人类1号染色体长臂(1q42.2),编码脯氨酸羟化酶2(PHD2)[3]。PHD2以氧浓度依赖的方式负调控HIF-1α。现已证实了EGLN1的独特表型在高原低氧适应中的重要性[4]。研究者发现藏族人的基因中有一种高频率的EGLN1变体(Asp4Glu/Cys127Ser),可降低低氧条件下促红细胞生成素的敏感性及红系祖细胞的增殖率[5]。因此,EGLN1与HIF介导的红系增生关系密切。基于此,本研究以HAPC模型大鼠为研究对象,探讨HAPC模型大鼠EGLN1的甲基化状态及与EGLN1 mRNA表达的关系,试图进一步揭示HAPC红系增生的机制。
1.材料与方法
1.1 动物
健康雄性SD大鼠,清洁级,6~8周龄,体重200±20 g,购自北京维通利华实验动物技术有限公司[许可证号:SCXK(京)2016-0006]。实验过程中对动物的处置符合2006年科学技术部发布的《关于善待实验动物的指导性意见》。所有大鼠均置于环境温度为18±2 ℃、相对湿度为40%~60%的环境中,随机喂食标准颗粒饲料和水。
1.2 HAPC模型大鼠制备方法
根据靳国恩等[6]提供的方法制备HAPC动物模型。将24只大鼠随机分为模型组和对照组,每组各12只,模型组在海拔4 300米的青海果洛玛多区饲养,对照组在海拔1 100米的甘肃天水麦积区饲养。根据2004年第六届国际高原医学和低氧生理学术大会制定的青海标准[7](血红蛋白>210g/L,红细胞压积>65%)判断是否造模成功。
1.3 取材方法
采用乌拉坦行腹腔麻醉(1g/kg)。分离腹主动脉,用采血针经腹主动脉采血于EDTA抗凝管中,除测血红蛋白、红细胞压积之外,另取2 mL血用于甲基化测定。剪开大鼠双侧股骨和胫骨,露出骨髓腔,用吸取1×PBS液的注射器反复冲洗,收集骨髓细胞悬液,离心(3000r/min)5 min,用300目滤网过滤,收集骨髓细胞。将收集好的骨髓细胞迅速冷冻于液氮中,进一步提取RNA。
1.4 实验方法
1.4.1EGLN1启动子区的甲基化状态检测
提取外周血基因组DNA,采用Zymo EZ DNA甲基化检测试剂盒对基因组DNA进行甲基化修饰,随后用吸附柱技术完成DNA的纯化并回收样本。所得DNA保存于-80 ℃冰箱。引物EGLN1pair1:上游序列为5′-GTAGTAGAATTTGGGATTTGG-3′,下游序列为5′-AACCTATTAATCACTTACTACCATCCA -3′;EGLN1pair2:上游序列为5′- AGTAAGYGTAAGGTTTTGGGTT-3′,下游序列为5′- CAACAAACAAAATACTTATTACAATTC-3′;EGLN1pair3:上游序列为5′-YGTTTAAGTAGTAGAGGTTGTAA-3′,下游序列为5′- CCTACTCTAATTAAAACTACAAAAA -3′。
PCR扩增体系(25μL):上、下游引物各0.5 μL,DNA甲基化转化溶液1.5 μL,Takara Ex Taq HS(RR006A)0.25 μL,10×Ex Taq Buffer 2.5 μL,dNTP Mixture(2.5mM each)2 μL,不足25 μL时用双蒸水补足。扩增反应条件:95 ℃预变性300 s;95 ℃ 15 s,58 ℃ 20 s,72 ℃ 20 s,共45 个循环;72 ℃延伸300 s 后冷却到4 ℃。将上述PCR产物通过1.5%琼脂糖凝胶电泳进行割胶回收,用GeneJET Gel Extraction Kit试剂盒纯化DNA片段,然后按比例混合不同的DNA片段,通过VAHTSTM Turbo DNA Library Prep Kit for Illumina试剂盒完成二代测序建库,最后通过IlluminaMiseq高通量测序平台进行测序。
1.4.2 大鼠骨髓EGLN1 mRNA水平检测
使用QIAGEN RNA simple Total RNA试剂盒从冷冻骨髓中提取总RNA。cDNA的合成采用Takara PrimeScriptTM RT Master Mix试剂盒。用TaKaRa TB GreenTM Premix Ex TaqTM试剂盒在7500 Real-time PCR仪(Bio-Rad,USA)上测定EGLN1 mRNA的表达水平。引物EGLN1pair1:上游序列为5′-AGGCAACGGGACAGGCTACG-3′,下游序列为5′-CGAAGAATACCTCCGCTCACCTTG-3′。引物GAPDH:上游序列为5′-AGTGCCAGCCTCGTCTCATA-3′,下游序列为5′-GAACTTGCCGTGGGTAGAGT-3′。基因相对表达量运用2-ΔΔCt法行数据分析。
1.4.3 统计学方法
采用SPSS17.0 统计软件行数据分析。数据用均数±标准差表示,两组间数据比较使用t检验。P<0.05为差异有统计学意义。
2.结果
2.1 模型组与对照组血红蛋白等指标的比较
表1显示,高原低氧HAPC组大鼠的血红蛋白和红细胞压积水平明显升高,与对照组相比差异有显著性意义(P<0.05)。

表1 两组间的生理指标
2.2 模型组与对照组大鼠EGLN1的甲基化状态分析
2.2.1 DNA提取
图1显示,采用1.0%琼脂糖凝胶电泳法检测DNA质量。DNA条带清晰,且无杂带和其他污染情况,说明DNA质量好,可以用于后续试验。

A:DNA samples of peripheral blood in HAPC model group.B:DNA samples of peripheral blood in control group.
2.2.2 甲基化分析区域说明
图2显示,EGLN1基因pair1、pair2及pair3引物的位置。
2.2.3EGLN1目标片段平均甲基化比例比较
表2显示,两组大鼠外周血EGLN1目标片段pair1、pair2和pair3的平均甲基化比例比较均无统计学差异(P>0.05)。
表2 两组间EGLN1目标片段平均甲基化比例
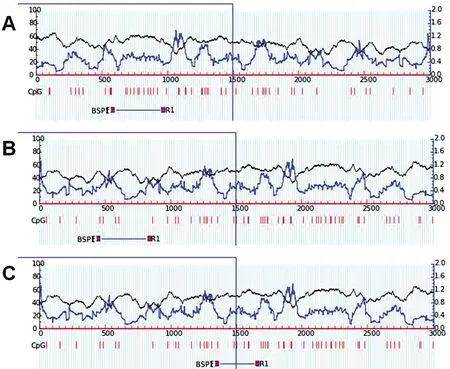
A:EGLN1 pair1;B:EGLN1 pair2;C:EGLN1 pair3
2.2.4 模型组与对照组大鼠骨髓EGLN1 mRNA测定
表3显示,模型组骨髓EGLN1 mRNA相对表达量较对照组低,但两组相比结果无统计学意义(P>0.05),与EGLN1甲基化结果相符。
表3 两组大鼠骨髓EGLN1 mRNA的相对表达量
3.讨论
多年来,研究者从红系信号转导异常、基因多态性及细胞的凋亡/增殖失衡等多方面研究HAPC的发病机制未能完全阐明其机理。DNA甲基化研究为揭示HAPC的发病机理提供了新的方向[2,8,9]。本研究应用亚硫酸氢盐测序法对HAPC组及对照组大鼠外周血EGLN1基因启动子区的甲基化状态进行检测,结果发现EGLN1基因pair1、pair2、pair3的甲基化状态及EGLN1 mRNA的表达在两组间均无差异,提示HAPC模型大鼠的发病与EGLN1的甲基化状态及基因表达无关。
PHD2由EGLN1编码而成,它是调节HIF稳定性的重要酶类。常氧下,HIF-1α亚基脯氨酸的羟基化过程可被PHD加速催化,肿瘤抑制蛋白(von Hipplelindau tumor suppressor protein,pVHL)能特异性地介导羟基化的HIF-1α经泛素化途径降解,从而抑制HIF-1α的功能[10]。低氧时,PHD羟基化作用减弱,HIF-1α的降解减少而在细胞中大量累积,促使HIF-1复合物与低氧反应元件(hypoxic response element,HRE)及辅助因子结合,最终调控HIF下游基因的转录增强了细胞对缺氧的适应[11]。既往对EGLN1的研究很多,Percy M 等[12]发现,由EGLN1突变导致的PHD2酶活性显著降低,与家族聚集性红细胞增多症发病相关。有研究者将大鼠EGLN1敲除,出现了红细胞数量显著增加和血管过度增生情况[13,14]。这些研究结果均表明EGLN1对红细胞增生具有重要意义。因此,充分了解EGLN1的结构和功能对开展HAPC研究至关重要。
HAPC发病率显著上升,除高原低氧因素外,还可能存在表观遗传学调控因素[2,9],但目前对于HAPC的表观遗传学研究非常有限。因此,我们首次对高海拔低氧环境HAPC模型大鼠的EGLN1甲基化状态与其发病的关系进行研究,虽然没有发现相关性,但是否存在表观遗传学的其他机制参与HAPC的发病仍有待进一步研究。
(致谢:感谢青海省果洛州玛多县人民医院在本研究中所给予的大力支持和帮助)
猜你喜欢
山东第一医科大学(山东省医学科学院)学报(2022年8期)2022-12-07
昆明医科大学学报(2022年1期)2022-02-28
世界科学技术-中医药现代化(2021年9期)2021-12-31
中国畜禽种业(2021年7期)2021-12-03
天津医科大学学报(2021年3期)2021-07-21
中学生物学(2020年10期)2020-12-25
中华老年口腔医学杂志(2019年2期)2019-04-28
安徽医学(2018年3期)2018-04-19
医学研究杂志(2015年12期)2015-06-10
军事体育学报(2015年2期)2015-02-27
