
SNAP29调节胃癌细胞顺铂耐药性的研究
2020-09-09 08:21张佳佳
安徽医科大学学报 2020年8期
张佳佳,喻 鑫,周 波,刘 弋
胃癌是消化系统常见恶性肿瘤之一,晚期胃癌预后较差。当前对于胃癌主要的治疗手段包括外科手术、放射治疗、化学药物治疗及靶向治疗。由于当前针对胃癌的靶向治疗药物较少,化学药物治疗仍然是晚期胃癌的重要治疗手段。顺铂是胃癌治疗的常用化疗药物,但药物耐受是亟待解决的重要问题[1-3]。该研究旨在检测SNAP29基因对胃癌细胞顺铂耐药性的作用,进一步分析SNAP29在胃癌患者中临床病理学意义。
1 材料与方法
1.1 病例资料本课题收集了安徽医科大学第一附属医院2017~2019年手术切除的患者石蜡组织标本,包括50例胃癌组织标本和50例正常胃组织标本,同时收集了胃癌患者的临床病理学参数,包括患者的年龄、性别、肿瘤大小、肿瘤淋巴结转移情况、肿瘤的分级和分期。患者对实验的开展知情同意。
1.2 细胞培养实验中涉及的胃癌细胞系MKN-45和AGS均来源ATCC (American Type Culture Collection),均根据推荐条件在37 ℃恒温5%CO2培养箱中进行培养,实验时选取处于对数生长期的细胞进行实验。
1.3 质粒合成和转染实验选取哺乳动物表达载体pIRESneo3 (Invitrogen)来构建SNAP29的过表达质粒。SNAP29的编码序列(GenBank识别码: NM_004782.4)克隆进pIRESneo3质粒,被命名为pIRESneo3-SNAP29。质粒转染实验使用lip2000 (QIAGEN)转染试剂,按照说明书推荐的条件进行,参考文献操作[4]。
1.4 Western blotWestern blot参考文献操作[4]。对转染后的细胞进行收集,使用细胞裂解液(Cell Signaling)裂解细胞并提取蛋白。提取的蛋白使用BCA Protein Assay kit (Pierce)试剂测定浓度。Western blot实验过程包括电泳、转膜、封闭、抗体孵育、酶标二抗孵育、显影过程。实验中主要的抗体包括兔SNAP29多克隆抗体(1 ∶1 000, 12704-1-AP)和鼠β-actin单克隆抗体(1 ∶10 000, 66009-1-Ig)(美国芝加哥Proteintech集团有限公司)。β-Actin作为内参。
1.5 MTT实验MTT实验参考文献操作[4]。具体过程如下:已转染的细胞24 h后消化打散成单细胞计数,种植于96孔细胞培养板中,每孔种植细胞5 000个,同组细胞种植3孔,细胞培养24 h后相应加入0、3、6 mg/L顺铂处理,48 h后进行MTT实验检测,测定570 nm的吸光度。
1.6 软琼脂克隆形成实验软琼脂克隆形成实验用于检测细胞克隆形成。具体过程是在6孔细胞培养板中铺设含0.5%琼脂糖的1640培养基(1.5 ml),凝固成下层胶,已转染的细胞24 h后消化打散成单细胞后计数,取相应数目的细胞加入含0.35%琼脂糖的1640培养基,种植于6孔板下层胶上,每孔对应5 000个细胞,1.5 ml培养基,室温凝固后,上层加入2 ml的1640完全培养基(含10%胎牛血清和1%青霉素链霉素双抗)。细胞种植48 h后相应加入0、6 mg/L顺铂处理48 h,细胞再培养4 d后检测克隆形成。每组重复3孔,进行统计分析。
1.7 免疫组织化学实验免疫组织化学实验参考文献操作[4]。具体过程包括石蜡组织切片的脱蜡、抗原修复、内源性过氧化物酶阻断、抗体孵育、辣根过氧化物酶标二抗孵育、DAB显色、苏木精复染、切片脱水封片等过程。实验使用迈新生物公司的Envision法免疫组织化学检测试剂盒和兔SNAP29多克隆抗体(1 ∶200, 12704-1-AP,美国芝加哥Proteintech集团有限公司)。SNAP29为胞质表达,在组织中细胞表达比例≥15%被定义为SNAP29阳性,<15%被定义为SNAP29阴性。
1.8 统计学处理采用SPSS16.0软件进行分析。本课题实验均独立重复3次,结果取平均值。MTT实验和软琼脂克隆形成实验结果使用t检验进行统计学分析。SNAP29表达水平与胃癌患者临床病理学参数相关性分析使用χ2检验。P<0.05为差异有统计学意义。
2 结果
2.1 在胃癌细胞MKN-45中过表达SNAP29检测细胞对顺铂的敏感性在本研究中构建了SNAP29过表达质粒pIRESneo3-SNAP29,转染MKN-45胃癌细胞系,记为MKN-45 SNAP29;对照pIRESneo3空质粒转染的MKN-45细胞系被记为MKN-45 Vec。Western blot实验显示MKN-45 SNAP29细胞中SNAP29蛋白表达水平高于MKN-45 Vec细胞(图1A)。MTT实验表明,MKN-45 SNAP29和MKN-45 Vec细胞在不加顺铂情况下的细胞活力没有显著差异,顺铂可以抑制MKN-45 SNAP29和MKN-45 Vec细胞的细胞活力;在加入3和6 mg/L顺铂情况下,MKN-45 SNAP29细胞的细胞活力高于MKN-45 Vec细胞(P<0.05)(图1B)。进一步细胞软琼脂克隆形成实验显示MKN-45 SNAP29和MKN-45 Vec细胞的克隆形成率无显著差异,顺铂抑制MKN-45 SNAP29和MKN-45 Vec细胞的克隆形成,但是在6 mg/L顺铂处理后,MKN-45 SNAP29细胞的克隆形成率高于MKN-45 Vec细胞(P=0.008 3)(图1C、D)。

图1 在胃癌细胞MKN-45中过表达SNAP29检测细胞对顺铂的敏感性
2.2 在胃癌细胞AGS中过表达SNAP29检测细胞对顺铂的敏感性AGS细胞转染SNAP29过表达质粒及对照空质粒,分别记为AGS SNAP29和AGS Vec。Western blot实验显示AGS SNAP29细胞中SNAP29蛋白表达水平高于AGS Vec细胞(图2A)。与MKN-45细胞系相似,AGS SNAP29和AGS Vec细胞在不加顺铂情况下的MTT细胞活力无显著差异,在加入3和6 mg/L顺铂后,AGS SNAP29和AGS Vec细胞的细胞活力均下降,但AGS SNAP29细胞的细胞活力高于AGS Vec细胞(P<0.05)(图2B)。细胞软琼脂克隆形成实验显示,在6 mg/L顺铂处理情况下,SNAP29过表达的AGS细胞软琼脂克隆形成率高于对照细胞(P=0.012 7)(图2C、D)。

图2 在胃癌细胞AGS中过表达SNAP29检测细胞对顺铂的敏感性
2.3 胃癌组织SNAP29的表达及其临床病理学参数相关性分析本课题组通过免疫组织化学实验检测了50例胃癌组织标本和50例正常胃组织标本中SNAP29的蛋白表达,结果显示在胃癌组织中SNAP29的阳性表达率是66.0%,在正常胃组织中SNAP29的阳性表达率是62.0%,两者差异无统计学意义(P>0.05)(表1、图3)。同时,本课题组收集了50例胃癌组织标本对应的患者临床病理学参数(包括患者的年龄、性别、肿瘤大小、肿瘤淋巴结转移情况、肿瘤的分级和分期),进一步对SNAP29表达情况与这些病理学参数进行了相关性分析(表2)。在胃癌患者中,SNAP29阳性表达与胃癌患者的肿瘤淋巴结转移(P=0.042)和肿瘤分期(P=0.023)具有相关性,而与胃癌患者的年龄、性别、肿瘤大小、肿瘤分级无相关性(P>0.05)。

表1 SNAP29在胃癌和正常组织中的表达

图3 SNAP29在胃癌组织和正常组织中的表达 ×200A:胃癌组织;B:正常组织
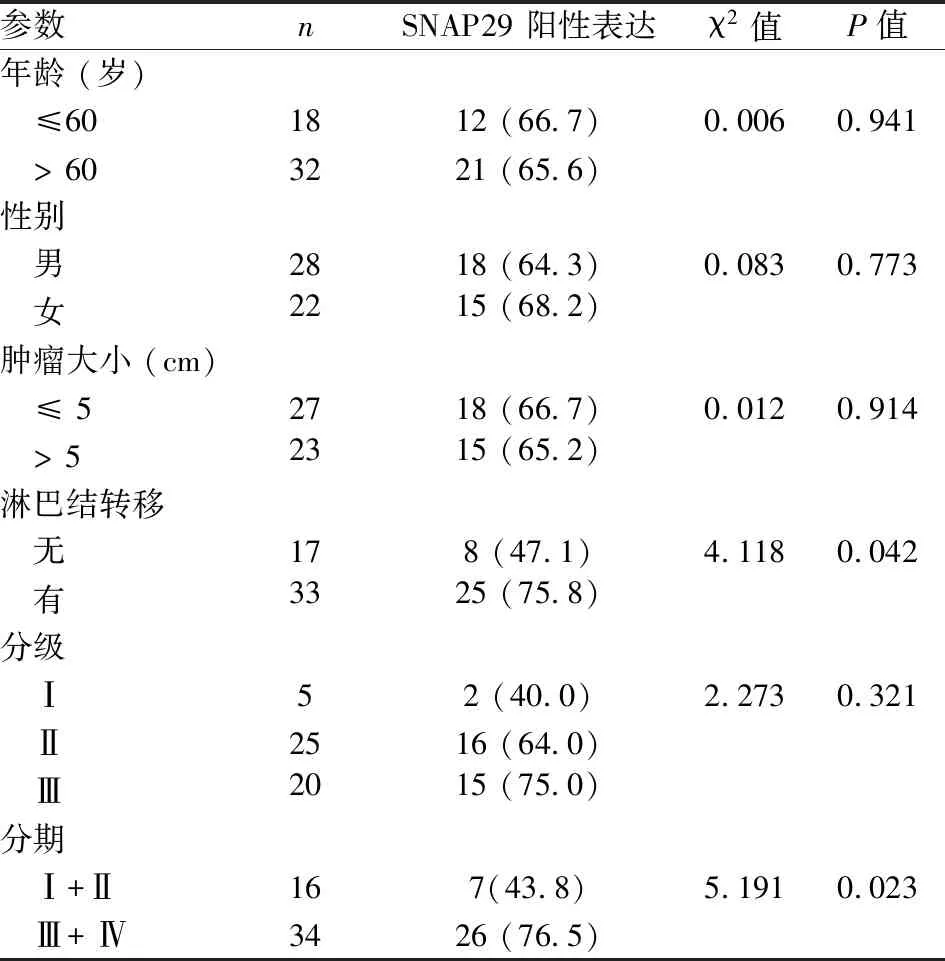
表2 SNAP29表达与胃癌患者临床病理参数相关性分析 [ n(%)]
3 讨论
胃癌是严重威胁人类健康的恶性肿瘤,早期胃癌预后相对较好,晚期胃癌治疗手段有限,预后较差。外科手术、辅助化疗或放疗是当前胃癌治疗的主要方式。针对胃癌常用靶向药物当前仅有曲妥珠单抗,它对HER2基因扩增的胃癌患者疗效较好,但HER2基因扩增在胃癌中的比例仅仅10%~15%[2-3,5]。顺铂是胃癌常用化疗药物之一,对晚期胃癌的治疗具有一定效果,但是长期用药后患者往往产生获得性药物耐受。针对胃癌的顺铂耐药,许多分子机制参与其中:Zhu et al[6]报道了miR-187靶向调控TGF-β/Smad信号通路参与调控胃癌细胞顺铂耐药;SLFN11的甲基化作用促进胃癌细胞的顺铂耐药性[7];METase/lncRNA HULC/FoxM1信号通路通过抑制细胞自噬从而降低胃癌顺铂耐药性[8]。关于胃癌顺铂耐药的相关研究不胜枚举,其相关机制复杂多样,更进一步的研究迫切而必要。本研究首次报道SNAP29参与促进胃癌细胞对顺铂的耐药,同时进一步揭示了SNAP29在胃癌患者中的病理学意义。
SNAP29属于突触体相关蛋白(SNAP)家族成员,参与SNARE(可溶性NSF附着受体)蛋白(SNARES)生物学功能。SNAP家族包括SNAP25、SNAP23和SNAP29,前两种蛋白分别是膜相关蛋白,控制突触传递,参与广泛的非神经元膜融合过程;SNAP29仅短暂的与膜结合,它包含一个酸性NPF基序,介导与内吞因子的结合[9]。许多研究[9-13]表明SNAP29在信号转导、细胞运动、细胞分裂、自噬和突触传递等过程中均发挥重要作用。Mastrodonato et al[10]预测SNAP29可能在肿瘤发生发展以及神经退行性变中发挥重要作用。但目前为止,关于SNAP29在人体肿瘤中研究的报道十分有限,关于SNAP29在人体胃癌中的功能仍然是未知的。本课题在胃癌细胞MKN-45和AGS中转染SNAP29过表达质粒,通过MTT实验和细胞软琼脂克隆形成实验研究SNAP29对细胞功能的作用。从图1和图2的结果来看,SNAP29的过表达对MKN-45和AGS细胞的细胞活力和细胞克隆形成能力均无显著作用,因此可以推断SNAP29对胃癌细胞的增殖能力并没有显著影响。
本课题在两种胃癌细胞系MKN-45和AGS中,通过MTT实验和细胞软琼脂克隆形成实验两种实验方法证明了SNAP29的过表达能够促进胃癌细胞对顺铂的耐药性。但限于实验条件限制,本课题没有揭示出SNAP29促进胃癌顺铂耐药的下游分子机制,这是下一步工作应该致力的方向。根据先前报道,SNAP29参与调控细胞自噬过程,SNAP29参与形成自噬体上的Syx17-Snap29复合物,是启动复合体与溶酶体上的Vamp8融合的关键[9,13-14]。因此在胃癌细胞中SNAP29很可能参与促进细胞自噬,从而介导了促进顺铂耐药的功能。Zhou et al[15]发现OGT的下调可以抑制SNAP29的O-GlcNAc修饰作用,从而促进了SNAP29-Stx17-VAMP8复合物的形成和细胞自噬过程,该机制促进了卵巢癌对于顺铂的耐药性。因此SNAP29在胃癌中促进顺铂耐药很可能是通过促进细胞自噬介导的,该具体机制在后续工作中将继续研究。
本课题检测了SNAP29在胃癌组织和正常胃组织中的蛋白水平,二者无显著差异。SNAP29在正常胃组织中的阳性率高达62.0%,这表明SNAP29在正常组织中也是发挥重要功能的。据报道[9-13]SNAP29参与细胞的信号转导、运动、有丝分裂、自噬和突触传递等过程,因此在胃组织中SNAP29很可能也参与了细胞的相关生理过程,但其具体作用仍是未知的,也是后续工作需要进一步研究的方向。胃癌患者病理学参数相关性分析结果显示,SNAP29与胃癌患者的年龄、性别、肿瘤大小和肿瘤分级无相关性,该结果证明了SNAP29在胃癌患者中的表达没有年龄和性别倾向,与肿瘤的增殖和生长非显著相关;SNAP29与胃癌患者的淋巴结转移和肿瘤分期具有相关性,该结果证明SNAP29可能对胃癌的转移具有一定促进功能。总体而言,SNAP29的高表达与胃癌患者的不良病理特征密切相关,在临床上具有一定意义。
总之,本课题首次报道了SNAP29在人体胃癌中的功能,明确了SNAP29在胃癌患者中的病理学意义。该研究对于进一步了解SNAP29的生理学功能,胃癌顺铂耐药机制具有一定意义,也为胃癌研究和治疗提供了新的靶标。
猜你喜欢
农产品加工(2022年17期)2022-10-17
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
化工进展(2022年7期)2022-08-01
新农业(2021年9期)2021-06-20
江西农业学报(2021年4期)2021-04-20
三农资讯半月报(2020年11期)2020-06-21
家禽科学(2020年3期)2020-05-13
长江丛刊(2017年17期)2017-11-25
医学研究杂志(2015年2期)2015-06-10
