
高效合成番茄红素酿酒酵母菌株的构建
2020-07-29 10:25孙玲王均华蒋玮李由然张梁丁重阳顾正华石贵阳徐沙
生物工程学报 2020年7期
孙玲,王均华,蒋玮,李由然,张梁,丁重阳,顾正华,石贵阳,徐沙
·工业生物技术·
高效合成番茄红素酿酒酵母菌株的构建
孙玲1,2,王均华1,2,蒋玮1,2,李由然1,2,张梁1,2,丁重阳1,2,顾正华1,2,石贵阳1,2,徐沙1,2
1 江南大学 粮食发酵工艺与技术国家工程实验室,江苏 无锡 214122 2 江南大学 生物工程学院,江苏 无锡 214122
番茄红素作为一种高附加价值的萜类化合物已受到国内外研究者的广泛关注。首先对酿酒酵母模式菌株S288c和YPH499合成番茄红素的能力进行分析比较,结果表明YPH499更适合作为底盘细胞用于番茄红素的合成。随后比较组成型启动子GPD、TEF1和诱导型启动子GAL1、GAL10对番茄红素合成的影响,结果发现以GPD、TEF1作为番茄红素合成途径基因、和的启动子,摇瓶发酵60 h后,番茄红素产量为15.31 mg/L;以GAL1和GAL10为启动子时,其产量为123.89 mg/L,提高8.09倍。继续改造甲羟戊酸(MVA) 途径,过量表达N-末端截短的关键酶基因t(3-羟基-3-甲基戊二酸单酰辅酶A还原酶),番茄红素产量为265.68 mg/L,单位菌体产量72.79 mg/g。文中所设计构建的异源表达番茄红素合成途径的酿酒酵母菌株单位细胞产量高,可以进一步改造和优化后用于番茄红素的工业化生产。
番茄红素,酿酒酵母,底盘细胞,诱导型启动子,单位菌体产量
番茄红素是一种深红色类胡萝卜素,其分子结构式为C40H56。作为人体必需的营养物质,番茄红素具有抗氧化性、提高免疫力、预防眼部及心血管疾病等功效,应用广泛。同时番茄红素作为一种很强的抗氧化剂,具有极强的清除自由基的能力,对防治肺癌[1]、胃癌[2]、前列腺癌[3]、卵巢癌[4]、结肠癌[5]等均有显著效果。目前,番茄红素主要由植物提取、化学合成和生物合成3种方法获得。从植物提取公众接受度高,但生长周期长、操作繁琐、收益率低且受环境影响较大;化学合成纯度较高,且不断优化合成路线后产业化比较稳定,但存在低产率及污染环境等问题[6];生物合成法虽会受到菌株遗传稳定性的影响,但因其具有低成本、生产周期短和绿色环保等优点,越来越受到人们的青睐。
酿酒酵母作为安全的模式微生物(GRAS),遗传可操作性强,在现代工业中已得到了广泛的应用[7-9]。酿酒酵母自身存在甲羟戊酸(MVA) 途径(图1),是番茄红素[10]、青蒿酸[11]等异源萜类物质合成的前体代谢途径。其中番茄红素以甲羟戊酸(MVA) 途径产生的IPP (异戊二烯基焦磷酸) 和DMAPP (二甲基丙烯焦磷酸) 作为前体,经由香叶基香叶基二磷酸合成酶(CrtE) 生成香叶基香叶基二磷酸酯(GGPP),在八氢番茄红素合成酶(CrtB) 作用下合成Phytoene (八氢番茄红素),之后由八氢番茄红素去饱和酶(CrtI) 作用产生一系列中间代谢产物,包括Phytofluene (六氢番茄红素)、ζ-Carotenet (ζ-胡萝卜素)、Neurosporene (链孢红素) 和终产物Lycopene (番茄红素)[12](图1)。
目前已有一些通过构建酿酒酵母细胞工厂进行番茄红素合成的策略[13-15]。浙江大学于洪巍团队[16]对番茄红素合成酶进行修饰,与来源于红发夫酵母的、以及酿酒酵母基因共表达,构建得到的菌株番茄红素单位细胞产量为24.41 mg/g DCW (1.61 g/L)。天津大学元英进团队[10]对番茄红素合成途径与宿主细胞进行修饰,筛选不同来源的基因以得到最佳组合,敲除酿酒酵母CEN.PK2菌株基因以增加胞质Acetyl-CoA前体供应,并敲除远端遗传基因、和及调控转录因子,重组菌在5 L发酵罐中单位细胞产量为55.56 mg/g DCW。武汉大学刘天罡团队[17]通过脂质工程与传统代谢工程结合,利用上调甲羟戊酸(MVA) 途径、增强胞质Acetyl-CoA前体供应以及促进辅因子NADPH合成等策略,构建的酿酒酵母产番茄红素单位细胞产量为56.2 mg/g。进一步通过增强三酰甘油(TAG) 代谢等方法,最终以酿酒酵母CEN.PK2菌株为底盘细胞构建得到工程菌,番茄红素单位细胞产量为70.5 mg/g DCW,通过高密度培养后达到73.3 mg/g DCW,此时番茄红素产量为2.37 g/L。本研究结合底盘细胞适配性和启动子筛选,利用CRISPR-Cas9技术构建得到高效合成番茄红素的酿酒酵母菌株,进一步改造和优化后可用于番茄红素的工业化生产。

图1 酿酒酵母合成番茄红素代谢路径
1 材料与方法
1.1 材料
1.1.1 菌株、质粒和培养基
本文涉及的所有质粒和菌株信息详见表1。大肠杆菌生长用SOB培养基(Super optimal broth):蛋白胨10 g/L,酵母粉5 g/L,氯化钠0.5 g/L,氯化钾0.186 g/L,六水合氯化镁2.207 g/L。酵母生长用YPD培养基:20 g/L葡萄糖、20 g/L胰蛋白胨、10 g/L酵母提取物。配置固体培养基时加入2.0%的琼脂粉。在菌株构建和摇瓶发酵过程中,培养基中添加的氨苄青霉素 (Ampicillin)、潮霉素(Hygromycin B)、硫酸诺尔斯菌素(Nourseothricin Sulfate,NTC)、遗传霉素(Geneticin,G418) 终浓度分别为100、500、100、500 µg/mL。YPG 培养基( 20 g/L半乳糖、20 g/L 胰蛋白胨,10 g /L 酵母提取物) 用于诱导表达Cre酶使G418抗性标记丢失。
1.1.2 引物
实验所用引物均由生工生物工程(上海) 股份有限公司合成,如表2所示。
1.1.3 主要工具酶和试剂
文中所用的2×PCR Master Mix、2×PCRMaster Mix购自杭州宝赛公司;Phanta®Max Super-Fidelity DNA polymerase购自南京诺唯赞生物科技有限公司;Fast DigestedTM快速限制性内切酶购自美国ThermoFisher Scientific公司;T4 DNA Ligase和pMD19T-simple 购自大连TaKaRa公司;硫酸诺尔斯菌素购自上海懋康生物科技有限公司;氨苄青霉素、PEG3350、醋酸锂、潮霉素、遗传霉素G418购自Sigma公司;质粒DNA提取试剂盒、DNA纯化试剂盒和DNA凝胶回收试剂盒购自北京博大泰克生物有限公司;番茄红素 (HPLC>=90%,CAS#502-65-8) 标准品购自上海源叶生物科技有限公司;鲑鱼精DNA购自北京索莱宝科技有限公司;蛋白胨、酵母粉、琼脂粉购自OXOID公司;三氯甲烷、丙酮为分析纯,甲醇为色谱纯,均购自国药集团化学试剂有限公司。
1.2 方法
1.2.1 番茄红素合成途径的构建
本实验所选取的表达番茄红素合成途径相关酶的编码基因来源情况如下:香叶基香叶基二磷酸合酶CrtE来源于曼地亚红豆杉(,Tm),八氢番茄红素合成酶CrtB来源于成团泛菌(,Pa),八氢番茄红素去饱和酶CrtI来源于三孢布拉氏霉菌(,Bt)。基因按照酿酒酵母密码子偏好性进行优化并规避常用酶切位点由苏州金唯智生物科技有限公司合成质粒PUC57-、PUC57-和PUC57-备用。

表1 质粒和菌株

表2 引物
Generated restriction enzyme sites are underlined and indicated in the table.
分别以质粒PUC57-、PUC57-和PUC57-为模板扩增、和片段。以本实验室保藏的酿酒酵母YPH499基因组为模板,扩增双向启动子GAL1GAL10及和两个基因的左右同源臂,并选择HⅠ作为和两个基因左右同源臂的酶切位点。分别以质粒pY26和PSH47-Hygr为模板扩增和终止子片段。将这些产物纯化后,通过融合PCR,将左同源臂与终止子片段融合,右同源臂与终止子片段融合、与启动子GAL1GAL10融合,根据凝胶电泳显示融合片段大小正确之后,将上述3组片段纯化再进行融合PCR,得到YJL064W-tCYC1-crtI-GAL1GAL10,最后通过TA克隆,连接至载体pMD19T-simple,得到质粒Ts-IB (GAL1GAL10)。用同样的方法构建Ts-IE (1GAL10)、Ts-IB(GPDTEF1)和Ts-IE (GPDTEF1)质粒,所有质粒送上海生工测序。
1.2.2 重组菌的筛选
本研究利用的CRISPR-Cas9基因编辑系统是一个双质粒系统,Cas9蛋白和向导gRNA分别由两个游离质粒在酿酒酵母菌株中表达以进行相应基因的敲除与敲入。转化酿酒酵母时,待转化目的物包括gRNA质粒和线性化的基因表达盒通过醋酸锂转化法转化至酿酒酵母,利用同源重组机制实现组装[18]。在固体培养基冷却至合适温度时,添加500 µg/mL G418用于敲除基因的转化子筛选,添加100 µg/mL NTC用于转化酿酒酵母Cas9质粒后转化子的筛选;同时添加 100 µg/mL NTC和500 µg/mL Hygromycin B用于转化酿酒酵母番茄红素合成途径表达盒后转化子的筛选。转化平板于30 ℃培养箱中倒置培养待长出转化子,经菌落颜色变化(合成番茄红素的菌株显深黄色或深红色) 和菌落PCR初步筛选,再通过基因组验证获得整合表达番茄红素工程菌株。
1.2.3 重组菌发酵实验
将平板活化得到的单克隆工程菌株接种于YPD液体培养基中,30 ℃、200 r/min振荡培养至对数生长中期(36 h左右),然后以600=0.5的初始菌体浓度转接于含50 mL YPD液体培养基的250 mL锥形瓶中,30 ℃、200 r/min振荡培养,定时取样对重组菌生长趋势、合成番茄红素水平、消耗葡萄糖情况以及生成副产物甘油、乙酸和乙醇情况进行测定。
1.2.4 番茄红素的提取
对发酵过程中提取的发酵液样品进行快速提取,具体方法如下所述:分别各取两等份1 mL发酵液,离心收集菌体,无菌水清洗后,其中一份菌体55 ℃烘干至恒重,称重计算菌体干重;另一份用于产物提取。向待提取番茄红素的离心管中加入等体积的玻璃珠(0.5 mm) 和1 mL氯仿,振荡破碎2 min,冰浴1 min,10 000 r/min离心 1 min,取上清液;黑暗条件下放置至细胞萃取完全后,离心取适量体积萃取液用氮吹仪将氯仿快速吹干,用丙酮复溶过0.22 µm有机膜后用超高效合相色谱(UPC2) 分析番茄红素产量。
1.2.5 番茄红素的含量测定
使用Waters超高效合相色谱仪(Ultra Performance Convergence Chromatography,UPC2)对番茄红素进行含量测定。检测条件为:配备PDA检测器的ACQUITY UPC2系统和专用色谱柱Viridis®HSS C18 SB Column 100A,1.8 µm;流动相为20%甲醇和80% CO2,流速1 mL/min,柱温40 ℃,检测波长450 nm,背压13.793 MPa (2 000 psi),进样量2 µL,重复3次测定;番茄红素标准品用于定量分析。将规格为20 mg的番茄红素标品用丙酮充分溶解,并加入0.05 g的2,6-二叔丁基-4-甲基苯酚(BHT) 作为保护剂[19],−70 ℃避光保存备用。并用丙酮稀释为不同浓度的标准样品后通过超高效合相色谱仪(UPC2) 测定以制作标准曲线。
1.2.6 胞外代谢物的检测
发酵液中葡萄糖、甘油、乙酸和乙醇的浓度利用高效液相色谱(赛默飞,UltiMate3000) 进行测定。以Bio-RadHPX-87H为色谱柱,5%的稀硫酸溶液作为流动相,流速为0.8 mL/min,柱温50 ℃,示差检测器。
2 结果与分析
2.1 番茄红素合成途径表达盒的构建
构建番茄红素合成途径表达盒如图2所示,将构建得到的质粒Ts-IB(GAL1GAL10) 和Ts-IE(GAL1GAL10) 分别用HⅠ酶切,结果如图3A所示,所得条带大小均与理论值4 722 bp/2 701 bp和4 735 bp/2 701 bp相符。同理,Ts-IB(GPDTEF1) 和Ts-IE(GPDTEF1) 用HⅠ酶切,凝胶电泳结果如图3B所示,所得条带大小均与理论值5 180 bp/2 701 bp和5 192 bp/ 2 701 bp相符,测序结果表明表达盒已成功构建。
已有研究表明作为一种转录因子,可抑制甲羟戊酸途径和麦角固醇合成酶;而基因的敲除可以提高红没药烯和类胡萝卜素的产量[20]。另外,刘天罡团队[17]研究发现可通过增加八氢番茄红素去饱和酶(CrtI) 基因拷贝数提高番茄红素的产量。根据以上分析,本研究利用CRISPR-Cas9技术在和两个基因的开放阅读框(ORF),分别同时敲入-和两组基因,实现基因的双拷贝和番茄红素合成途径的异源表达。
2.2 不同底盘细胞对番茄红素途径表达水平的影响
异源路径与底盘细胞的适配性是高效利用微生物作为细胞工厂的关键因素之一[10]。本研究以酿酒酵母菌株S288c与YPH499为底盘细胞,在和两个基因开放阅读框(ORF) 分别同时敲入tCYC-crtI-GPDTEF1-crtB-tADH和tCYC1-crtI-GPDTEF1-crtE-tADH1两组表达盒,得到酿酒酵母工程菌株S288c-CrtE/B/I和YPH499-CrtE/B/I。对重组菌进行摇瓶发酵实验,提取发酵液样品,按照上述条件用UPC2进行定性和定量分析。结果如图4所示,两株菌的生长能力与番茄红素合成能力均有较大差异(图4)。发酵60 h,YPH499-CrtE/B/I菌体生长能力明显低于S288c-CrtE/B/I (图4A)。酿酒酵母S288c- CrtE/B/I在发酵60 h后番茄红素产量为4.11 mg/L,YPH499-CrtE/B/I番茄红素产量为15.31 mg/L,是前者的3.7倍,说明酿酒酵母YPH499虽然较模式菌株S288c菌体生长能力低,但产物积累能力更强,因此更适合作为底盘细胞合成番茄红素。
2.3 比较启动子对番茄红素途径表达水平的影响
本研究进一步比较启动子对控制番茄红素合成途径表达的影响。首先敲除基因使半乳糖诱导型启动子GAL1和GAL10实现组成型表达[21]。在此基础上,分别转化组成型启动子GPD和TEF1和半乳糖诱导型启动子GAL1和GAL10控制番茄红素合成途径表达盒,获得YPH499GAL80-CrtE/B/I和YPH499GAL80-Galpr- CrtE/B/I两株酿酒酵母工程菌。摇瓶发酵3 d,结果如图5所示,两株菌生长速率差别不大(图5A),但合成番茄红素的水平明显存在巨大差异(图5B)。当以GAL1和GAL10控制番茄红素合成途径表达时,番茄红素产量在发酵60 h即可达到123.89 mg/L,产量提高8.09倍。如图5C所示,4株工程菌发酵48 h后,发酵液颜色即呈现明显差别。相比其他3株菌发酵液呈现黄色,YPH499GAL80-Galpr- CrtE/B/I发酵液呈现出明显的红色。结果表明以YPH499菌株作为底盘细胞,半乳糖诱导型启动子GAL1和GAL10控制表达番茄红素合成途径效果更佳。对YPH499GAL80-Galpr-CrtE/B/I胞外代谢物包括发酵液中葡萄糖、甘油、乙酸、乙醇的浓度进行测定(图5D),结果表明,发酵液中葡萄在12 h–30 h之间被快速消耗用于菌体生长,整个过程基本没有乙酸产生,但存在一定量的副产物乙醇和少量的甘油。
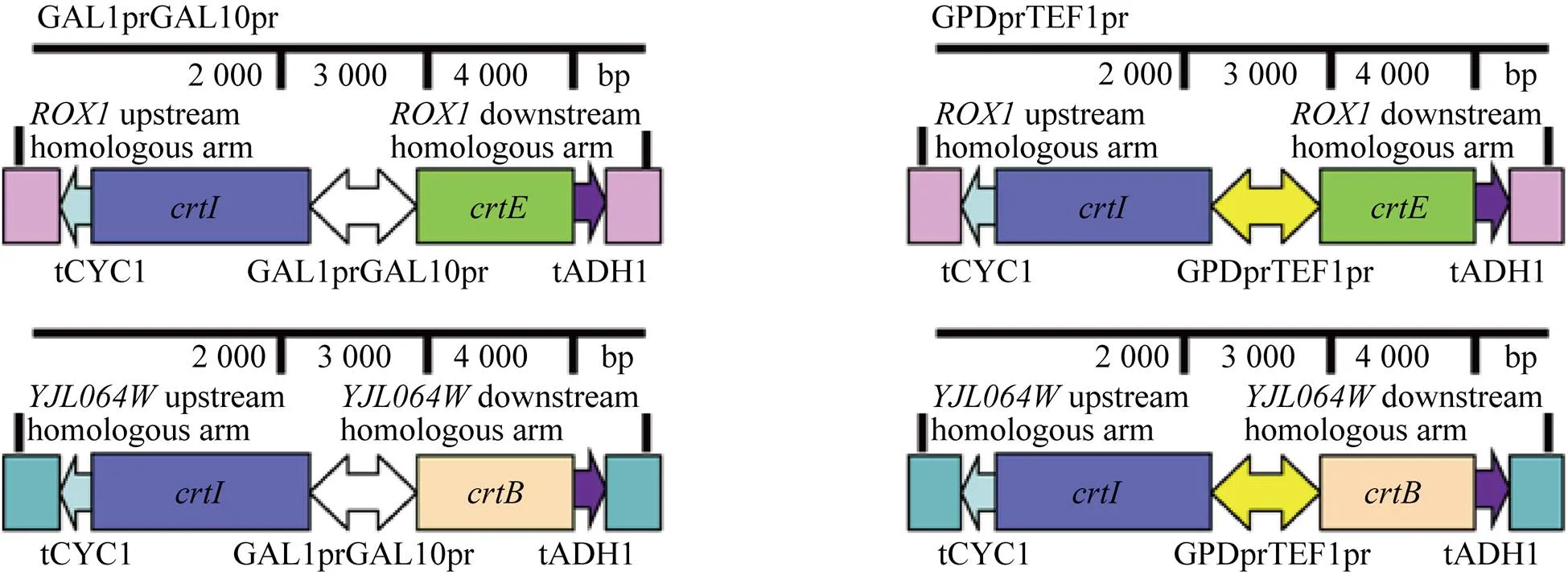
图2 番茄红素合成途径表达盒

图3 番茄红素合成途径表达盒的酶切验证
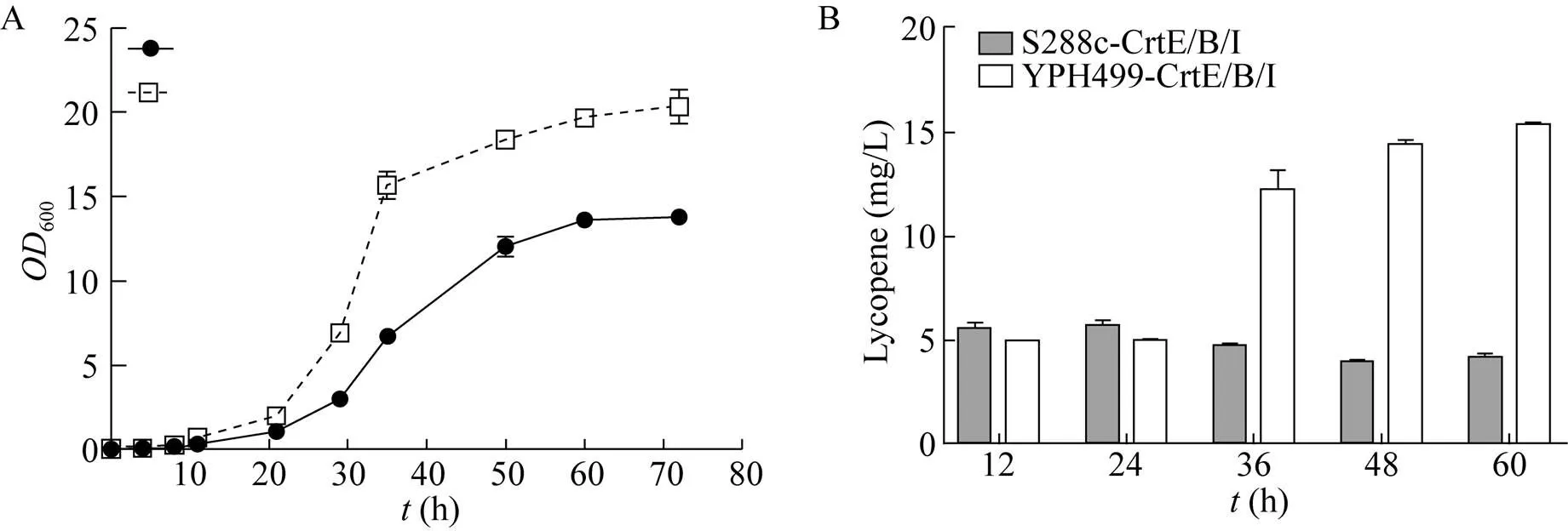
图4 底盘细胞对重组菌合成番茄红素的影响(A:菌体生长曲线(OD600);B:工程菌摇瓶发酵生产番茄红素)
图5 启动子对重组菌合成番茄红素的影响(A:不同启动子条件下的菌体生长曲线;B:工程菌摇瓶发酵生产番茄红素;C:番茄红素在不同菌株中的积累;D:发酵过程中的胞外代谢产物和葡萄糖浓度变化曲线)
Fig. 5 Effect of promoters on synthesis of lycopene by engineered S. cerevisiae. (A) The effect of different promoters on cell growth. (B) Lycopene production in Shake flask fermentation using engineered S.cerevisiae. (C) Accumulation of lycopene in different strains. (D) The concentration of extracellular metabolites including glucose concentration, glycerol concentration, and ethanol concentration in the fermentation of YPH499GAL80-Galpr-CrtE/B/I.
2.4 过量表达tHMG1对番茄红素合成能力的影响
为了进一步提高番茄红素产量,以本实验室构建的一株将甲羟戊酸(MVA) 途径基因全部上调的重组菌株WHtE作为宿主细胞(数据未发表)异源表达番茄红素合成途径。结果如图6所示,摇瓶发酵60 h,番茄红素的产量并没有显著提高,仅为12.34 mg/L,且菌株生长受到严重抑制,推测可能基因改造过多对菌株生长造成影响反而不利于产物合成。文献报道t作为甲羟戊酸(MVA) 途径的关键酶可以有效解除麦角固醇的反馈抑制[22]。如图6B所示,当只过表达甲羟戊酸(MVA) 途径的关键酶基因t时,最终番茄红素产量达到265.68 mg/L。此时,单位菌体番茄红素产量达到72.79 mg/g,而菌体干重仅为3.65 g/L。
3 讨论
本研究发现,选择合适的底盘细胞能够极大地提高产物合成效率。选择酿酒酵母YPH499作为底盘细胞,仅表达外源番茄红素合成途径,番茄红素产量达到15.31 mg/L,是S288c的3.7倍。李霞等[23]分别以酿酒酵母 INVSc1和CEN.PK2-1C为底盘细胞,发酵120 h,番茄红素产量分别为0.025 mg/L和0.052 mg/L,说明不同遗传背景酿酒酵母的番茄红素合成能力不同,与本研究结果相符。同时,启动子作为启动基因转录调控的元件,其活性强弱直接影响基因的表达水平,进而影响代谢途径转化效率和目标产物产量[24]。本研究结果表明在双向启动子调控下,番茄红素产量达到123.89 mg/L,相比组成型强启动子调控的菌株产量提高8.09倍。现有研究表明,在敲除的前提下,启动子虽然不受半乳糖的调控,但是其转录强度随着葡萄糖浓度的降低而提高[10]。在发酵过程中,前期由于启动子活性较低,番茄红素积累量较低,随着葡萄糖的消耗,启动子的转录强度逐渐提高,产物大量积累。
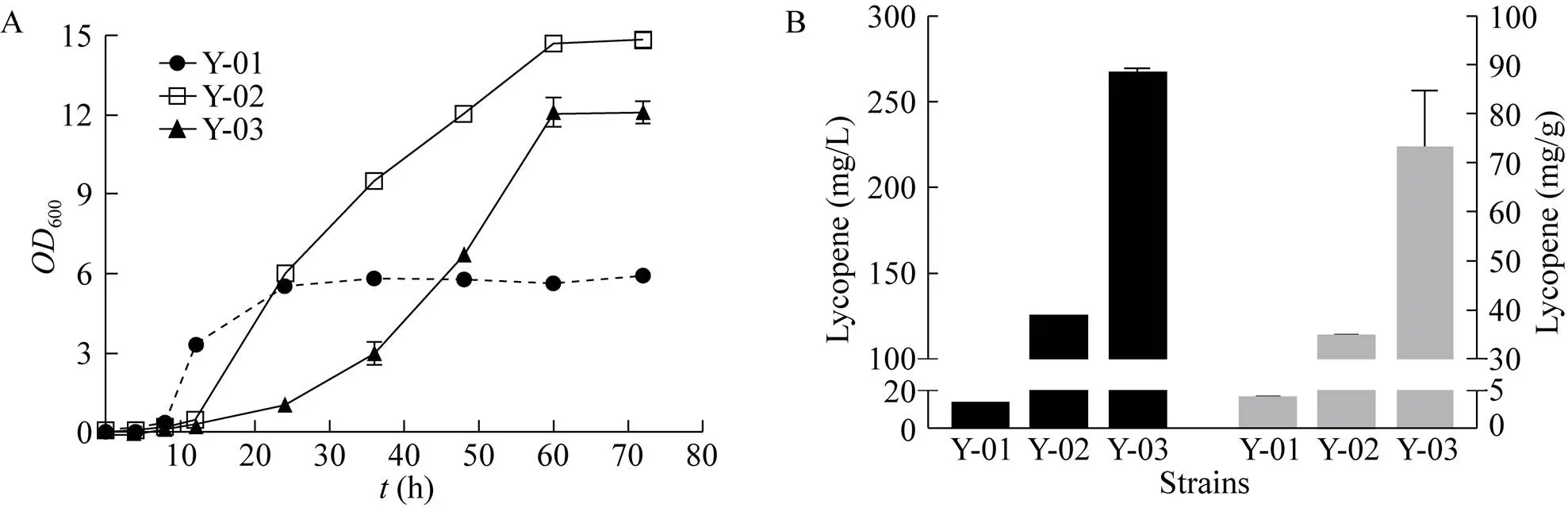
图6 调控MVA途径对重组菌合成番茄红素的影响
为了进一步增强番茄红素合成途径的代谢通量,过表达甲羟戊酸途径的关键酶调控基因,最终番茄红素产量达到265.68 mg/L,单位菌体产量为72.79 mg/g DCW。经过改造,单位菌体产量已经达到较高水平,但产量仍然较低的原因是由于发酵结束菌体干重仅为3.65 g/L。由于番茄红素积累在微生物胞内,不会主动分泌到培养基中,因此单位细胞产量是一个重要指标。后续的改造重点在于增加菌体积累,通过流加碳源、优化培养基组分、碳氮比等方法进行高密度培养[10]。综上所述,本研究构建了一株番茄红素单位菌体产量较高的菌株,后续将对发酵条件进行优化,对重组菌进行高密度培养,提高细胞浓度,以实现番茄红素的高产。
[1] Rakic JM,Liu C,Veeramachaneni S,et al. Lycopene inhibits smoke-induced chronic obstructive pulmonary disease and lung carcinogenesis by modulating reverse cholesterol transport in ferretsCancer Prev Res, 2019, 12(7): 421–431.
[2] Han H, Lim JW, Kim H. Lycopene inhibits activation of epidermal growth factor receptor and expression of Cyclooxygenase-2 in gastric cancer cellsNutrients, 2019, 11(9): 2113.
[3] Li DB, Chen LN, Zhao WR, et al. MicroRNA-let-7f-1 is induced by lycopene and inhibits cell proliferation and triggers apoptosis in prostate cancerMol Med Rep, 2016, 13(3): 2708–2714.
[4] Xu J, Li YF, Hu HY,et al. Effects of lycopene on ovarian cancer cell line SKOV3: Suppressed proliferation and enhanced apoptosisMol Cell Probes, 2019, 46: 101419.
[5] Langner E, Lemieszek MK, Rzeski W,et al. Lycopene, sulforaphane, quercetin, and curcumin applied together show improved antiproliferative potential in colon cancer cellsJ Food Biochem, 2019, 43(4): e12802.
[6] Clinton S. Lycopene: Chemistry, biology, and implications for human health and diseaseNutr Rev, 1998, 56(2): 35–51.
[7] Dai ZB, Liu Y, Zhang XN, et al. Metabolic engineering offor production of ginsenosidesMetab Eng, 2013, 20: 146–156.
[8] Dai ZB, Liu Y, Huang LQ, et al. Production of miltiradiene by metabolically engineeredBiotechnol Bioeng, 2012, 109(11): 2845–2853.
[9] Han JY, Seo SH, Song JM, et al. High-level recombinant production of squalene using selectedstrainsJ Ind Microbiol Biotechnol, 2018, 45(4): 239–251.
[10] Chen Y, Xiao WH, Wang Y, et al. Lycopene overproduction inthrough combining pathway engineering with host engineeringMicrob Cell Fact, 2016, 15: 113.
[11] Zhang CB, Liu JJ, Zhao FL, et al. Production of sesquiterpenoid zerumbone from metabolic engineeredMetab Eng, 2018, 49: 28–35.
[12] Cunningham XF Jr, Sun Z, Chamovitz D, et al. Molecular structure and enzymatic function of lycopene cyclase from thesp.strain PCC7942Plant Cell, 1994, 6(8): 1107–1121.
[13] Yamano S, Ishii T, Nakagawa M, et al. Metabolic engineering for production ofβ-carotene and lycopene inBiosci Biotechnol Biochem, 1994, 58(6): 1112–1114.
[14] Shi B, Ma T, Liu TL, et al. Systematic metabolic engineering offor lycopene overproductionJ Agric Food Chem, 2019, 67(40): 11148–11157.
[15] Bahieldin A, Gadalla NO, Al-Garni SM, et al.Efficient production of lycopene inby expression of syntheticgenes from a plasmid harboring thepromoterPlasmid, 2014, 72: 18–28.
[16] Xie WP, Lv XM, Ye LD, et al. Construction of lycopene-overproducingby combining directed evolution and metabolic engineeringMetab Eng, 2015, 30: 69–78.
[17] Ma T, Shi B, Ye ZL,et al. Lipid engineering combined with systematic metabolic engineering offor high-yield production of lycopeneMetab Eng, 2019, 52: 134–142.
[18] Gietz RD, Woods RA. Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol methodMethods Enzymol, 2002, 350: 87–96.
[19] Xu F, Yuan QP, Dong HR. Determination of lycopene andβ-carotene by high-performance liquid chromatography using sudan I as internal standardJ Chromatogr B, 2006, 838(1): 44–49.
[20] Özaydin B, Burd H, Lee TS,et al. Carotenoid-based phenotypic screen of the yeast deletion collection reveals new genes with roles in isoprenoid productionMetab Eng, 2013, 15: 174–183.
[21] Peng BY, Wood RJ, Nielsen LK,et al. An Expanded Heterologous GAL Promoter Collection for Diauxie-Inducible Expression inACS Synth Biol, 2018, 7(2): 748–751.
[22] Donald KA, HamptonRY, Fritz IB. Effects of Overproduction of the Catalytic Domain of 3-Hydroxy-3-Methylglutaryl Coenzyme A Reductase on Squalene Synthesis inAppl Environ Microbiol, 1997, 63(9): 3341–3344.
[23] Li X,Zhao JY, Lou XD, et alOptimization of the adaptability ofproducing lycopeneFood Fermentat Ind, 2018, 44(6): 24–29 (in Chinese).李霞,赵金雨,娄兴丹,等.酿酒酵母合成番茄红素的适配性优化.食品与发酵工业, 2018, 44(6):24–29.
[24] Keasling JD. Synthetic biology and the development of tools for metabolic engineeringMetab Eng, 2012, 14(3): 189–195.
Construction of a highly efficient synthetic lycopene engineered
Ling Sun1,2, Junhua Wang1,2, Wei Jiang1,2, Youran Li1,2, Liang Zhang1,2, Zhongyang Ding1,2, Zhenghua Gu1,2, Guiyang Shi1,2, and Sha Xu1,2
1,,214122,,2,,214122,,
Lycopene, as a high value-added terpene compound, has been widely concerned by researchers at home and abroad. Firstly, the ability of lycopene synthesis ofmodel strains S288c and YPH499 was analyzed and compared. The results showed that YPH499 was more suitable for lycopene synthesis as yeast chassis. Subsequently, the effects of constitutive promotersGPD,TEF1and inducible promotersGAL1,GAL10on Lycopene synthesis were compared. The results showed that whenGPDandTEF1were used as promoters of,andin lycopene synthesis pathway, the production of lycopene was 15.31 mg/L after 60 h fermentation in shaking flask. WhenGAL1andGAL10were used as promoters, the production was 123.89 mg/L, which was 8.09 times higher. In addition, the methylvaleric acid (MVA) pathway was further modified to overexpress the key enzyme gene of N-terminal truncation, t(3-hydroxy-3-methylglutaryl coenzyme A reductase). The lycopene production was 265.68 mg/L, and the yield per cell was 72.79 mg/g. Thestrain designed and constructed in this study can express lycopene in high yield per cell, thus could be used in the industrial production of lycopene after further construction and optimization.
lycopene,, yeast chassis, inducible promoter,yield per cell
10.13345/j.cjb.190524
November 23, 2019;
April10, 2020
Supported by:National Natural Science Foundation of China (No. 31571817).
Sha Xu. Tel/Fax: +86-510-85914371; E-mail: xusha1984@jiangnan.edu.cn
国家自然科学基金 (No. 31571817) 资助。
2020-05-15
http://kns.cnki.net/kcms/detail/11.1998.Q.20200514.1529.003.html
孙玲, 王均华, 蒋玮, 等. 高效合成番茄红素酿酒酵母菌株的构建. 生物工程学报, 2020, 36(7): 1334–1345.
Sun L, Wang JH, Jiang W, et al. Construction of a highly efficient synthetic lycopene engineered. Chin J Biotech, 2020, 36(7): 1334–1345.
(本文责编 郝丽芳)
猜你喜欢
中国调味品(2022年10期)2022-10-13
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
当代水产(2022年1期)2022-04-26
食品安全导刊(2021年20期)2021-08-30
江西农业学报(2021年4期)2021-04-20
热带农业科学(2020年7期)2020-08-31
三农资讯半月报(2020年11期)2020-06-21
食品界(2019年2期)2019-03-10
家庭医药·快乐养生(2018年12期)2018-12-28
