
1株NADC30-like重组猪繁殖与呼吸综合征病毒的分离鉴定及遗传变异分析
2020-05-11 03:35邓紫艳董雅琴吴发兴李晓成
中国动物传染病学报 2020年3期
邓紫艳,张 锋,董雅琴,吴发兴,段 纲,李晓成
(1.中国动物卫生与流行病学中心,青岛 266032;2.云南农业大学动物医学院,昆明 650201)
猪繁殖与呼吸综合征(porcine reproductive and respiratory syndrome,PRRS)是一种严重危害全球养猪业的重要经济性传染病,其病原为猪繁殖与呼吸综合征病毒(Porcine reproductive and respiratory syndrome virus,PRRSV)。PRRS的主要临床症状为母猪繁殖障碍和各年龄段猪呼吸道疾病[1]。该病于1987年在美国首次被报道,其后迅速传播至许多养猪国家。2006年,我国暴发的高致病性PRRS(Highly PRRS,HP-PRRS)给养猪业造成了巨大的经济损失[2]。由于PRRSV具有广泛的变异和毒株多样性,导致新毒株不断出现。2014年,我国出现了以NADC30-like为优势病毒株的流行病毒株,造成许多规模性猪场发生疫情[3]。PRRSV是有囊膜、单股正链RNA病毒。根据基因组核苷酸序列及抗原性差异,PRRSV分为2个基因型,欧洲型(1型),代表病毒株为Lelystab virus(LV),和北美洲型(2型),代表病毒株为VR2332[4]。PRRSV基因组全长约15 Kb,编码至少10个开放阅读框(open reading frame,ORF),主要包括ORF1a、ORF1b、ORF2a、ORF2b、ORF3~5、ORF5a和ORF6-7[5-6]。基因组5'端的ORF1a和ORF1b编码2个长的多聚蛋白pp1a和pp1ab。多聚蛋白被切割成至少12个非结构蛋白,在病毒的复制和免疫调控中发挥重要作用[7]。ORF2a、ORF2b、ORF3和ORF4分别编码病毒的次要结构蛋白GP2a、E、GP3和GP4,ORF5、ORF6和ORF7分别编码病毒的主要结构蛋白GP5、M和N,通过与宿主蛋白相互作用调控病毒转录[8-9]。病毒中和表位位于结构蛋白,可以诱导中和抗体的产生。我国猪场出现的NADC30-like病毒株,与HPPRRSV的Nsp2基因存在30个不连续氨基酸缺失不同,NADC30-like病毒株的Nsp2出现了131个不连续氨基酸缺失的分子特征。有研究表明,当前正在广泛使用的商业化疫苗对NADC30-like病毒株仅能提供有限的免疫保护作用[10]。为进一步了解山东地区NADC30-like PRRSV流行病毒株的遗传变异情况,本研究从山东威海某疑似PRRS发病规模化猪场发病死亡猪肺脏病料中分离获得1株PRRSV毒株,经全基因组测序鉴定和分析,该毒株为NADC30-like,将其命名为SDwh株。经生物信息学软件SimPlot分析发现,其基因组存在重组现象。本研究中对SDwh株遗传演化的分析新型重组病毒对于PRRSV监测和防控具有重要意义。
1 材料与方法
1.1 病料和主要试剂 病料采自山东省威海市某疑似暴发PRRS猪场送检的发病保育仔猪肺脏组织,置于-20℃冰箱冷藏备用。病毒DNA/RNA提取试剂盒和DH5α感受态细胞购自北京天根生化科技有限公司;反转录酶(M-MLV)、RNA酶抑制剂和dNTP购自Promega公司;DL2000 DNA marker、胶回收试剂盒购自宝生物工程(大连)有限公司;KOD FX Neo高保真DNA聚合酶购自东洋纺(上海)生物科技有限公司;pJET1.2/blunt Cloning Vector购自ThermoFisher公司;DMEM和胎牛血清购自Invitrogen公司;Marc-145细胞由中国动物卫生与流行病学中心畜病监测室保存。
1.2 引物设计与合成 根据GenBank中VR2332、NADC30等PRRSV代表性毒株全基因组序列,设计了10对特异性引物用于扩增其全基因组序列(表1),引物由北京华大基因生物技术有限公司合成。

表1 PRRSV全基因组扩增用引物Table1 Primers used for amplification of PRRSV complete genome
1.3 病料样品处理 将采集的发病猪肺组织剪碎,匀浆研磨后用灭菌PBS按1∶5的比例稀释制成悬液,反复冻融3次,10 000×g离心10 min,取上清液经0.22 μm微孔滤膜除菌,滤液保存于-80℃。
1.4 病毒分离培养和鉴定 Marc-145细胞用含10%胎牛血清的DMEM培养传代,至6孔细胞培养板中长满单层时,接种含病毒的滤液200 μL,置于37℃、5%CO2细胞培养箱孵育1 h,弃上清液,加入2 mL含5%FBS的DMEM维持液,置于37℃、5%CO2细胞培养箱培养3~5 d。每日观察细胞病变效应(cytopathic effect,CPE)。如果出现CPE,取200 μL阳性培养物再传3代,至80%细胞出现CPE时收毒。取分离传代后的第3代病毒上清液提取核酸,进行常见猪源病毒(猪瘟病毒、伪狂犬病毒、猪圆环病毒、猪流行性腹泻病毒和猪细小病毒)的特异性PCR鉴定。同时利用抗PRRSV N蛋白特异性单克隆抗体对分离病毒进行间接免疫荧光特异性鉴定。
1.5 全基因组扩增和测序 取收获的第3代上清液提取病毒RNA,使用随机引物反转录制备cDNA。PCR反应条件:94℃预变性2 min,98℃变性10 s、56℃退火30 s、68℃延伸2 min,30个循环;72℃延伸7 min。PCR扩增产物经1%琼脂糖凝胶电泳鉴定,按照胶回收试剂盒说明回收纯化后,连接至pJET1.2/blunt克隆载体上,转化DH5α感受态细胞,挑菌鉴定后,送至北京华大基因生物技术有限公司测序。
1.6 全基因组序列比对与分析 利用Lasergene的SeqMan软件对全基因组序列进行拼接,用MegAlign与国内外分离毒株进行同源性比较。采用Mega 6.0软件进行序列分析,使用邻位相连法(Neighborjoining,NJ)构建遗传进化树。PRRSV代表毒株基因组序列均来自GenBank。
1.7 全基因组重组分析 利用SimPlot v3.5.1生物信息学软件,分析分离病毒株的重组情况,滑动窗口为500 bp,滑动步伐为20 bp。
2 结果
2.1 病料样品的RT-PCR检测 以提取的病料样品RNA为模板,使用PRRSV特异性引物ORF5F和ORF5R PCR扩增ORF5。结果显示,扩增出的目的片段大小为510 bp,与预期一致。
2.2 病毒分离培养和鉴定 病料接种Marc-145细胞,36 h后产生明显的CPE变化,细胞成灶状变圆突起、膨大,60 h后细胞皱缩、脱落(图1)。PCR鉴定结果显示,分离病毒中能够扩增出510 bp的PRRSV特异性目的片段,而其他常见猪源病毒检测结果均呈阴性。同时IFA检测结果显示,分离病毒感染Marc-145细胞中出现特异性的荧光(图2)。

图1 病毒在Marc-145细胞上的病变结果(200×)Fig.1 Cytopathic effect of virus in Marc-145 cell (200×)
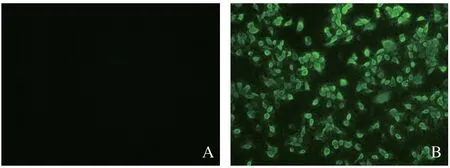
图2 分离病毒IFA特异性鉴定Fig.2 Identification of isolated virus by immunofluorescence assay (IFA)
2.3 全基因组扩增和测定 利用10对特异性引物对分离病毒基因组进行分段扩增,经琼脂糖凝胶电泳检测显示,扩增的10个片段大小与预期一致(图3)。扩增片段胶回收后,连接至pJET1.2/blun克隆载体上,由北京华大基因生物技术有限公司测序。利用SeqMan软件将测序结果拼接,获得PRRSV全基因组序列,长度为15 010 bp,命名为SDwh株。

图3 PRRSV全基因组扩增结果Fig.3 The whole genome amplification of PRRSV
2.4 SDwh全基因组同源性分析 将SDwh株与从GenBank中下载的国内外代表病毒株序列比对显示,SDwh株与北美病毒株NADC30的同源性最高,为93.1%;与我国分离的NADC30-like病毒株CHsx1401、JL580的同源性分别为91.1%和88.4%;与我国HP-PRRSV JXA1、HuN4株及疫苗病毒株JXA1-P80同源性分别为85.3%、85.3%和85.2%;与北美代表病毒株VR2332、中国PRRSV经典病毒株CH-1a及疫苗病毒株CH-1R的同源性均为85.3%;与欧洲型代表病毒株Lelystad virus同源性最低,仅为60.6%。表明SDwh株与北美病毒株NADC30亲缘关系最近,与我国之前流行的HP-PRRSV和经典PRRSV亲缘关系较远。
2.5 SDwh株全基因组遗传演化分析 使用Mega 6.0软件构建遗传进化树,结果显示,PRRSV毒株分为两个群,以LV为代表的欧洲型和以VR2332为代表的美洲型。我国2型PRRSV又分为4个谱系(lineage 1、lineage 3、lineage 5和lineage 8),SDwh株与美国病毒株NADC30和中国的NADC30-like病毒株同属于lineage 1(图4)。表明SDwh株是北美病毒株NADC30进入我国后逐渐演化产生的新的病毒株。进一步分析发现,NADC30-like PRRSV亚群中病毒株又被分为不同分支,其中山东地区2016年分离病毒株SDYG1606和SDlz1601进化关系较近,2017年分离病毒株SD17-36和SD17-38进化关系较近,而SDwh毒株与上述病毒株进化关系较远。
2.6 SDwh株Nsp1、Nsp2和GP5氨基酸序列比对 将SDwh株与国内外PRRSV代表病毒株序列进行核苷酸和氨基酸同源性分析。结果显示,与其他病毒株相比,SDwh株与NADC30在Nsp2~ORF7编码区具有更高的相似性,分别为89.0%~97.9%和85.6%~100%;而在Nsp1编码区域,SDwh株与JXA1-P显示出更高的核苷酸和氨基酸同源性,分别为96.8%和95.6%(表2),提示该毒株可能存在重组现象。

图4 基于PRRSV全基因组的遗传演化分析Fig.4 Phylogenetic analysis of PRRSV whole genome
Nsp2氨基酸比对结果显示,与VR2332和CH-1a相比,SDwh株Nsp2具有不连续131个氨基酸的缺失,分别位于在322~432位(111个)、483位(1个)和504~522位(19个),同样的缺失情况出现于美国分离的病毒株NADC30和我国近年来流行的NADC30-like(如CHsx1401和JL580)中。同时在584~585位缺失2个氨基酸(图5A)。GP5氨基酸比对结果显示,与美洲型代表病毒株VR2332相比,SDwh株GP5在非中和表位1(27~30位氨基酸)存在V27A和A29V的突变,在非中和表位2(180~197位氨基酸)存在V185A、I189V、R191K和V192I的变异,非中和表位的突变与NADC30和NADC30-like病毒株JL580、CHsx1401相似。中和表位(37~45位氨基酸)无氨基酸突变(图5B)。表明SDwh Nsp2和GP5不仅具有NADC30-like病毒株的特征,还表现出新的氨基酸缺失变异。
2.7 SDwh株的重组分析 为了进一步分析SDwh株的重组情况,利用SimPlot生物信息学软件对分离病毒SDwh株基因组进行相似性比对分析,滑动窗口为500 bp,滑动步伐为20 bp。相似性图谱结果显示,SDwh株存在重组现象,重组位点位于Nsp2中(2066 nt),该断点将SDwh株基因组分为2个区域(A和B),区域A与JXA1-P80亲缘关系最近,区域B与NADC30(2008年美国分离的2型PRRSV代表病毒株)亲缘关系最近(图6)。表明SDwh株是美国病毒株NADC30进入国内后与JXA1-P80重组产生的新型病毒。

表2 SDwh株与其他代表性毒株PRRSV基因组不同区域的核苷酸和氨基酸同源性比较Table 2 Nucleotide and amino acid identities of different regions of SDwh genome with other representative PRRSV strains
3 讨论
PRRSV作为一种迅速变异和演化的RNA病毒,其持续变异和毒株多样性加剧了该病预防和控制难度。自我国1996年首次分离到PRRSV毒株CH-1a以来,该病逐渐在猪场流行。2006年出现和流行的HP-PRRSV导致的疫情给我国养猪业造成了巨大的经济损失。近年来发现,NADC30-like新病毒株在我国流行,成为当前的优势流行病毒株之一。为了解山东地区PRRSV流行病毒株的遗传变异情况,本研究从山东省威海地区发病猪场采集阳性病料,并成功分离到PRRSV毒株SDwh。对该毒株进行全基因组测序和序列分析,结果显示,SDwh株与美国高毒力病毒株NADC30高度同源;与VR2332相比,SDwh Nsp2编码区存在131个氨基酸不连续缺失,GP5蛋白抗原表位存在6个位点的明显突变,符合NADC30-like PRRSV特有的氨基酸特征。同时SDwh株Nsp2在其他位置出现2个氨基酸的缺失,Nsp2氨基酸变化可能导致其抗原性发生改变。
依据SDwh全基因组核苷酸序列构建进化树,遗传演化分析表明,SDwh属于NADC30-like PRRSV亚群。进一步分析编码蛋白发现,SDwh Nsp1和JXA1-P80 Nsp1具有更高的氨基酸相似性,提示SDwh可能存在重组现象。有研究表明NADC30-like病毒株具有高致病性,易与我国已有的HP-PRRSV发生毒株间的重组,临床上正在使用的商业化疫苗对该类毒株缺乏免疫保护作用[10]。重组作为PRRSV遗传演化的一种模式,Li等[11]报道PRRSV毒株Em2007是田间流行株HP-PRRSV WUH1和疫苗病毒株CH-1R的重组病毒,动物实验显示,Em2007的致病性高于CH-1R亲本毒株CH-1a,表明重组在产生新毒株的同时,也存在致病性增强的情况发生。重组分析发现,SDwh可能是北美毒株NADC30和疫苗毒株JXA1-P80重组产生的新毒株,目前猪场使用的疫苗主要是经典PRRSV或HP-PRRSV传代致弱后的弱毒疫苗,与SDwh株的亲缘关系较远,因此不能为猪提供有效保护,导致猪场出现PRRSV感染发病情况发生。作为同期在PAM上分离的SD17-38毒株,其在Marc-145细胞未见明显CPE,动物实验显示其对猪具有高致病性[12],而SDwh株可以同时在PAM和Marc-145细胞中产生明显细胞病变,推测SDwh株具有更强的适应能力,为其在猪群中的传播提供更有利的条件,导致该类型毒株在山东地区猪群中长期存在。

图5 PRRSV Nsp2(A)和GP5(B)氨基酸序列比对Fig.5 Sequence alignment of PRRSV Nsp2 (A) and GP5 (B) amino acid
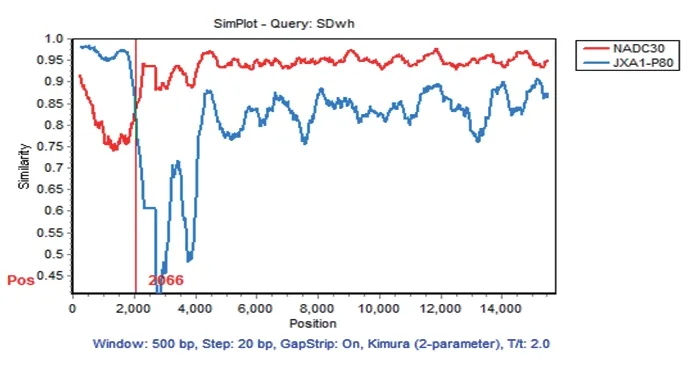
图6 PRRSV SDwh株全基因组重组分析Fig.6 Recombination analysis of the whole genome of PRRSV SDwh strain
NADC30-like病毒株在我国的流行导致PRRSV的病毒株多样性增加,其变异速度快、易重组的特性,极易导致疫病的局部暴发和流行强度增强,从而增加疫情控制难度。新型PRRSV疫苗的开发变得尤为重要,通过对山东地区流行的NADC30-like病毒株的分离及遗传演化研究,为流行病毒株的重组特性分析以及新型PRRSV毒株疫苗的研制提供一定的数据支撑。
猜你喜欢
动物医学进展(2022年9期)2022-11-26
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
今日农业(2021年17期)2021-10-14
养猪(2021年2期)2021-04-12
今日农业(2020年17期)2020-12-15
西部论丛(2019年31期)2019-10-14
中国动物保健(2019年4期)2019-08-06
