
滤泡辅助性T细胞在实验性免疫性不育小鼠中的表达
2020-04-07 10:46曹晓丹魏任雄李如瑶贝华锋
中华临床免疫和变态反应杂志 2020年1期
曹晓丹,魏任雄,李如瑶,贝华锋
男性不育病因复杂,其中8%~10%与免疫因素有关。免疫性不育是在某些外因或内因的诱发下,自身免疫耐受状态被打破,持续迁延的自身免疫对自身抗原产生异常的免疫应答,导致免疫调节失衡,以致疾病的发生。早期研究认为细胞免疫Thl/Th2平衡的变化及Th细胞辅助B细胞介导的体液免疫在免疫性不育疾病的发生中扮演重要角色[1-2]。但近来研究发现辅助B细胞完成体液免疫应答的Th2细胞仅起到调节作用,而真正辅助B细胞产生体液免疫应答的是一群新的CD4+Th细胞亚群——滤泡辅助性T(T follicular helper,Tfh)细胞,Tfh细胞的发现被认为是Th-B细胞相互协作研究的重大突破[3-4]。多项研究证实在多种自身免疫性疾病中均存在Tfh细胞数量增加或功能异常,是自身免疫性疾病免疫紊乱的关键因素[5-8]。而Tfh细胞在实验性免疫性不育小鼠中的表达特征目前尚不清楚。本研究通过建立免疫性不育小鼠动物模型,研究免疫性不育小鼠脾脏淋巴细胞中Tfh细胞比例的变化及其与B淋巴细胞水平之间的关系。
1 材料与方法
1.1 材料
1.1.1 实验动物
无特定病原(SPF)级BALB/c小鼠,雄性,35只,6~8 周龄,体重16~18 g,购自上海斯莱克实验动物有限责任公司,生产许可证号:SCXK(沪)2017-0005。对BALB/c小鼠昼夜12 h 交替光照,自由食水,于室温 (23±1)℃条件下饲养。动物实验符合浙江中医药大学动物实验研究中心实验动物福利伦理要求。
1.1.2 仪器和试剂
流式细胞计数仪 (美国BD 公司);细胞计数仪 (美国Bio-Rad公司);APC标记抗小鼠CD4单克隆抗体、PerCP/Cy5.5标记抗小鼠CXC趋化因子受体5(CXCR5)单克隆抗体、FITC标记抗小鼠可诱导共刺激分子(ICOS)单克隆抗体、PEcy7标记抗小鼠CD19单克隆抗体、PE标记抗小鼠CD3单克隆抗体 (美国eBioscience公司);小鼠抗精子抗体检测试剂盒(上海恒远生物科技有限公司)
1.2 方法
1.2.1 免疫性不育小鼠模型的建立
每次免疫前取10周龄雄性BALB/c小鼠5只,共3次,CO2安乐死后取双侧睾丸组织,用生理盐水冲洗,置于M199获能液中,剪碎,37 ℃静置15 min,尼龙网(孔径200 μm)过滤,37 ℃培育1.5 h,调精子数为2.0×106/mL,加入完全福氏佐剂,研磨混匀作为抗原。
取雄性BALB/c小鼠20只,随机分为正常组10只、实验组10只。实验组双侧腹股沟颈背部脊柱两侧皮下多点注射0.5 mL抗原作为首次免疫,10 d后于上述相同部位加强免疫。正常组注射生理盐水。分别于初免前及每次加强免疫后眼眶静脉采血,分离血清,ELISA检测血清抗精子抗体(antisperm antibody, AsAb)。第3次加强免疫后检测小鼠血清抗精子抗体水平,观察免疫性不育小鼠阳性只数,筛选成功造模的小鼠进行后续研究[9]。
1.2.2 小鼠脾脏淋巴细胞的分离
小鼠经CO2安乐死后,用75%的酒精浸泡2~3 min,减少毛发所造成的污染。将小鼠移入超净工作台内,暴露并切开腹腔壁膜,用镊子夹住脾脏,并剪去与其相连的组织。在培养皿中加入适量的PBS溶液,放置细胞筛,将脾脏取出放于细胞筛中。取干净的注射器,用其按压处的末端对组织进行碾压。经过细胞筛之后,细胞悬浮于培养皿溶液中。洗涤细胞筛,收集细胞筛外PBS至50 mL离心管中,400 g离心15 min;去上清,加红细胞裂解液2 mL,重悬。 PBS洗涤后重悬,除去其中不可溶的组织纤维,悬浮液取10 μL细胞计数,调整细胞密度为1×106/mL。
1.2.3 流式细胞术检测小鼠脾淋巴细胞内Tfh细胞、B细胞的比例
上述方法分离获得的脾淋巴细胞取1×106个细胞,PBS洗涤2次,4 ℃,1 000 r/min离心5 min。取100 μL PBS重悬细胞,分别加入APC标记抗小鼠CD4单克隆抗体、PerCP/Cy5.5标记抗小鼠CXCR5单克隆抗体、FITC标记抗小鼠ICOS单克隆抗体,或PEcy7标记抗小鼠CD19单克隆抗体、PE标记抗小鼠CD3单克隆抗体各5 μL,混匀后4 ℃避光孵育30 min。4 ℃,1 000 r/min离心5 min,弃上清,PBS洗涤后重悬细胞,用PBS稀释至300~500 μL,流式细胞仪检测[10]。
将分离的脾脏淋巴细胞进行多色免疫荧光标记,以CD4+CXCR5+ICOS+作为Tfh细胞的标志进行分析。先以CD4+设门,分析CD4+细胞中CXCR5+ICOS+T细胞的比例。以CD3-CD19+作为B细胞的标志进行分析。
1.3 统计学分析
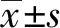
2 结果
2.1 免疫性不育小鼠模型的建立
对第3次加强免疫后的小鼠血清进行AsAb检测,结果显示,实验组小鼠血清吸光度值为(0.45±0.04),对照组小鼠血清吸光度值为(0.18±0.01)。实验组10只小鼠AsAb吸光度/正常对照组≥2.1,即为AsAb(+),表明实验组10只BALB/c小鼠均造模成功。
2.2 对照组和实验组Tfh 细胞表达的差异
免疫性不育小鼠脾脏淋巴细胞中CD4+CXCR5+ICOS+T 细胞比例(4.15±1.03)% ,明显高于健康对照组(1.05±0.25)%(n=10,t= 10.15,P<0.000 1)(图1)。
2.3 对照组和实验组B细胞表达的差异
免疫性不育小鼠脾脏淋巴细胞中CD3-CD19+B 细胞比例(8.05±1.97)%,明显高于健康对照组(2.81±0.47)% (n=10,t=8.44,P<0.000 1)(图2)。
2.4 免疫性不育小鼠Tfh 细胞比例与B 细胞比例相关性分析
免疫性不育小鼠脾脏淋巴细胞中Tfh 细胞与B 细胞比例相关性分析结果显示,免疫性不育小鼠Tfh 细胞与B 细胞比例呈正相关(r=0.58,P<0.05)。
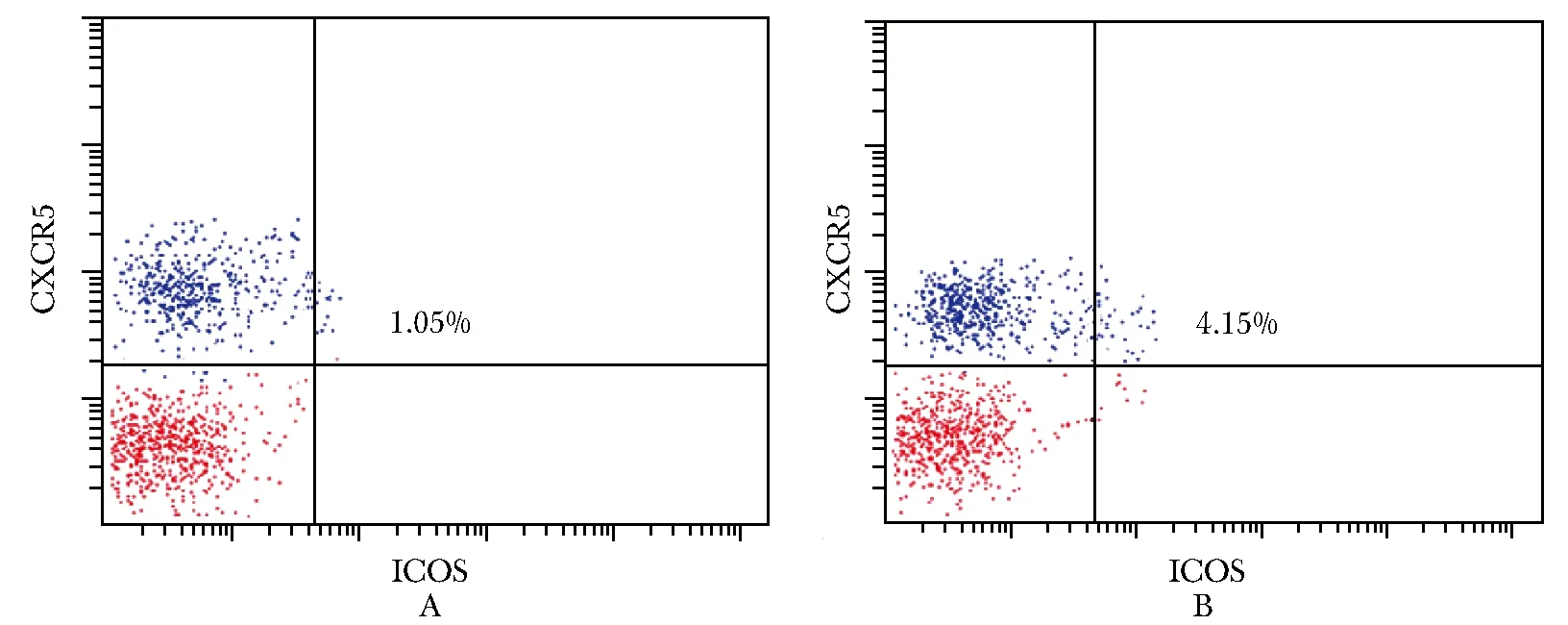
图1小鼠脾脏淋巴细胞中CD4+CXCR5+ICOS+T细胞比例(n=10)
Fig1Proportion of CD4+CXCR5+ICOS+T cells in spleen lymphocytes of mice(n=10)
A:正常对照小鼠;B:免疫性不育小鼠
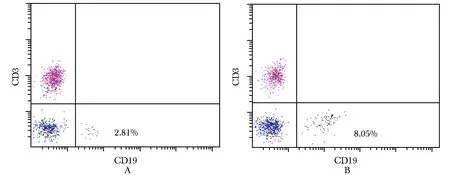
图2小鼠脾脏淋巴细胞中CD3-CD19+B细胞比例(n=10)
Fig2Proportion of CD3-CD19+B cells in spleen lymphocytes of mice(n=10)
A:正常对照小鼠;B:免疫性不育小鼠
3 讨论
男性免疫性不育是由于输精管手术、睾丸损伤、生殖道感染等造成血睾屏障以及生殖道免疫屏障的破坏而使精子受到免疫攻击,从而产生自身抗体的一种自身免疫性疾病[12]。T、B淋巴细胞免疫功能异常被认为是免疫性不育发病机制的关键因素,Th细胞辅助B细胞增殖分化为浆细胞和记忆性B细胞,完成抗体类别转换和抗体亲和力成熟;当T、B淋巴细胞的自我限制和自我耐受机制出现异常时,会引起机体针对自身抗原的免疫反应,Th细胞及相关细胞因子高表达,导致B细胞功能亢进,产生抗精子抗体。AsAb是免疫性不育机体中的复杂病理产物,AsAb制动精子,阻碍精子和卵子结合而致不育不孕,参与自身免疫性疾病的发生和发展。
Tfh细胞是由IL-21等诱导Th0分化产生,是真正意义上辅助B细胞的一群新的CD4+Th细胞亚群[13-14]。Tfh细胞定位于淋巴滤泡,高表达趋化因子受体CXCR5及共刺激分子ICOS,并通过表达的CD40配体(CD40L)和ICOS与B细胞表达的CD40和ICOS配体(ICOSL)相互作用,辅助B细胞在生发中心的存活、增殖,促进B细胞向浆细胞分化、抗体类别转换,在自身免疫性疾病中扮演重要角色。Tfh细胞持续高表达的趋化因子受体CXCR5,作为其迁移定位的重要转运分子,与其配体CXC趋化因子配体13(CXCLl3)结合刺激作用下迁移到B淋巴细胞滤泡中进行分化成熟,形成生发中心[15-16]。Tfh细胞在生发中心的形成过程中扮演重要角色。Tfh细胞敲除的小鼠生发中心形成缺陷,B细胞发育受阻,抗体产生水平下降[17]。相反,一些自身免疫疾病小鼠Tfh细胞比例增加,造成生发中心过度增生,甚至出现异位生发中心,并诱导B细胞的过度活化及自身抗体的过度分泌[18-19]。正常情况下,生发中心内经过体细胞高频突变的自身反应性B细胞会被可溶性的抗原诱导凋亡,但在自身免疫性疾病中B细胞的选择机制紊乱以及在识别相同抗原的Tfh细胞的辅助下,发生Tfh-B相互作用,高亲和力B淋巴细胞增殖、分化为记忆性B淋巴细胞或浆细胞,促进免疫球蛋白类别的转换,导致某类高滴度自身抗体的产生,机体体液免疫应答发生紊乱。提示Tfh 细胞功能正常与否对自身免疫性疾病机体体液免疫应答起着关键作用[20-21]。
Tfh细胞与多种自身免疫性疾病如类风湿关节炎、系统性红斑狼疮、重症肌无力、自身免疫性肝炎、原发性干燥综合征等密切相关。Ma 等[5]发现类风湿关节炎患者外周血Tfh表达数量及IL-21表达水平均较健康对照组明显升高,Tfh 细胞可促进细胞因子IL-21的分泌,IL-21与IL-21R结合后能够促进Tfh和B细胞的生长、分化,促进抗体产生和类别转换。系统性红斑狼疮患者外周血Tfh细胞比例高于健康对照组,且与疾病活动度、血清总IgG、抗核抗体滴度及抗ds-DNA均呈正相关[7],其发病机制可能与Tfh 过度表达、IL-21过量分泌及高滴度自身抗体的产生有关[8, 22]。在其疾病小鼠模型中,阻断IL-21及ICOS-ICOSL 信号通路可以引起致病性自身抗体的减少,阻止终末器官的病理学改变[23]。重症肌无力患者外周血CD4+CXCR5+T 淋巴细胞比例、IL-21 水平明显高于健康对照者,且与疾病严重程度相关,B 淋巴细胞上IL-21R与IL-21 相互作用,促进B 淋巴细胞增殖、免疫球蛋白类别的转换,诱导患者血清中抗体向IgGl 和IgG3 亚型转换[24-25],Tfh 密切参与了重症肌无力的发病过程。在免疫性不育疾病中,有研究显示免疫性不育患者外周血Tfh细胞及IL-21含量均较健康对照组显著升高,Tfh细胞水平与抗精子抗体的产生有显著相关性,提示Tfh细胞可能与产生自身抗体的异常体液免疫应答有关联[26]。
众多的研究提示Tfh细胞在多种由自身抗体介导的小鼠自身免疫性疾病模型和人类自身免疫性疾病中起着重要作用,大多数自身免疫性疾病中都伴随有Tfh细胞及B淋巴细胞形成过度。本次实验结果发现,免疫性不育小鼠脾脏淋巴细胞中CD4+CXCR5+ICOS+Tfh细胞和CD3-CD19+B细胞较健康对照组显著升高,且Tfh 细胞与B 细胞呈显著正相关。本研究探讨了Tfh 细胞在免疫性不育小鼠中的表达特征,为进一步研究Tfh 细胞在免疫性不育疾病中的具体作用及机制奠定了基础。
猜你喜欢
首都食品与医药(2022年19期)2022-11-19
现代临床医学(2022年1期)2022-02-12
昆明医科大学学报(2021年4期)2021-07-23
昆明医科大学学报(2021年2期)2021-03-29
天津医科大学学报(2021年2期)2021-03-29
中华养生保健(2020年9期)2021-01-18
保健与生活(2020年5期)2020-03-20
保健文汇(2020年12期)2020-02-02
中国现代药物应用(2019年14期)2019-08-16
医药前沿(2019年7期)2019-01-05
