
miR-150及其靶基因IRAK2在变应性鼻炎小鼠模型中的表达
2020-04-07 10:46郭照萌曾宪海刘丽芳蒋赢坤邓祖辉邬汉成刘志强刘江琦
中华临床免疫和变态反应杂志 2020年1期
郭照萌,曾宪海,刘丽芳,蒋赢坤,邓祖辉,邬汉成,刘志强,刘江琦
变应性鼻炎(allergic rhinitis,AR)是一种由特异性IgE介导的I 型变态反应[1],主要以阵发性喷嚏、清水涕、鼻塞、鼻痒等鼻过敏症状为特征,严重影响人类的生活质量[2]。目前,AR 的病因及发病机制尚未明确,现代医学认为固有免疫系统和适应性免疫系统失衡以及环境因素、遗传因素在AR发病过程中扮演重要角色[3]。
MicroRNA(miR)是一类非编码单链小分子RNA,能够与特定的mRNA结合,通过与靶基因mRNA 3′非翻译区(3′UTR)的完全或不完全配对[4],降解靶基因mRNA或抑制其翻译,从而对蛋白编码基因进行转录后调控,参与个体发育、细胞凋亡、增殖及分化等生命活动[5]。近年来研究发现在过敏性疾病模型中,多种miR的表达异常,且与体内的Th1/Th2免疫应答紊乱密切相关。miR-150作为miR家族中一员,与多种免疫细胞的分化和发育有关。miR-150通过特异性结合多个下游靶基因参与多种信号通路的调控,参与了多种临床疾病的发生发展过程[6]。本研究通过检测miR-150及其靶基因IRAK2在变应性鼻炎小鼠模型中的表达,旨在表明miR-150及其靶基因IRAK2在AR中的作用及意义。
1 资料与方法
1.1 实验材料
5~6周龄雄性BALB/c小鼠购自广东省实验动物中心[动物使用许可证号SCXK(粤)2018-0002,合格证号No.44007200060985]。小鼠在空调动物房[温度:(25±1)℃,湿度:(65±5)%]适应性饲养一周后开始实验。本研究使用的所有实验方案均经深圳市耳鼻咽喉研究所动物实验伦理委员会批准(中国深圳);无菌饲料(广东省实验动物中心);鸡卵清蛋白(美国,Sigma-Aldrich,A5253);氢氧化铝粉(分析纯,天津市大茂化学试剂厂); OVA-sIgE、 IL-4和IL-13 ELISA试剂盒(R&D公司),IRAK2抗体(ABCAM公司);RNA提取试剂盒(Invitrogen公司);cDNA反转录试剂盒(Thermo Scientific公司);miR-150实时定量PCR试剂盒(吉玛基因);引物合成购于生工生物工程(上海)股份有限公司。
1.2 模型建立
将小鼠随机分为2组,每组10只。变应性鼻炎模型组(AR)使用卵清蛋白致敏,将100 μg卵清蛋白+2 mg氢氧化铝溶于0.1 mL生理盐水中,分别于造模的第0、7、14天对模型组小鼠进行腹腔注射,第15天起至第28天每日分别用500 μg卵清蛋白溶于10 μL生理盐水中,对小鼠进行滴鼻,每个鼻孔10 μL,连续激发14 d。正常对照组(NC)同期使用生理盐水替代。
1.3 模型评价
通过激发,AR组小鼠会出现打喷嚏、挠鼻、抓脸、鼻溢等症状,末次激发后观察30 min,通过观察其相关症状进行变应性鼻炎动物行为学评分。评分标准:1分,鼻痒,轻摩擦鼻几次;2分,抓挠鼻面部不止;3分,到处摩擦。喷嚏:1~3个为1分,4~10个为2分,11个以上为3分;清涕:流到前鼻孔为1分,超过前鼻孔为2分,流涕满面为3分。以叠加法记录总分,总分超过5分即为建模成功。
1.4 组织病理学分析
所有小鼠均于末次激发后24 h内处死,其中每组4只小鼠取出完整鼻腔固定于4%的多聚甲醛中24 h,根据文献[7]配制适量醋盐酸甲醛脱钙液,脱钙6 h,之后置入5%甲酸中脱钙4周,然后包埋成蜡块。蜡块制作完成后,行石蜡切片,采取4 μm厚度切片。使用HE染色,镜检。镜下可见AR组小鼠鼻甲黏膜肿胀,嗜酸性粒细胞浸润,对照组未见明显黏膜肿胀及嗜酸性粒细胞浸润,说明造模成功。
1.5 外周血OVA-sIgE、 IL-4和IL-13检测
小鼠于末次激发24 h,颈椎脱臼处死后,摘取眼球取其外周血,室温静置30 min后,离心收集上清,-20 ℃保存。用ELISA 检测两组小鼠血清OVA-sIgE、 IL-4和IL-13的浓度(pg/mL),具体步骤参照ELISA试剂盒说明书进行。
1.6 实时定量PCR
取每组剩余6只小鼠鼻黏膜,应用RNeasy Mini Kit试剂盒提取小鼠鼻黏膜组织总RNA,紫外分光光度计测定RNA的浓度及纯度;根据 TaKaRa PrimeScriPt Ⅱ1st Strand cDNA Synthesis kit试剂盒说明书将2 μg RNA反转录为cDNA,反应终体系为20 μL;使用miR-150荧光定量PCR试剂盒和TaKaRa SYBR Premix Ex Taq Ⅱ在中ABI 7500进行RT-PCR扩增,95 ℃ 3 min预变性,随后95 ℃ 12 s和62 ℃ 40 s,40个循环采集荧光信号,并使用(2-△△Ct)方法进行测定。引物序列见表1。
1.7 Western blot检测IRAK2蛋白表达
将小鼠鼻黏膜组织剪碎后放入预冷的匀浆器中(每组n=4),加入含有蛋白酶和磷酸酶抑制剂的裂解液进行研磨,研磨后冰上裂解15 min,离心收集上清,-20 ℃保存备用。根据BCA蛋白定量法测定蛋白浓度,蛋白变性后进行10% SDS-PAGE电泳分离蛋白,电转至PVDF膜上,3% BSA封闭1 h,加入IRAK2(1∶1 000)单克隆抗体(购自英国abcam),4 ℃孵育过夜,TBST洗涤3次,每次10 min;加入HRP标记的二抗,室温震荡孵育1 h;按上述洗涤方法清洗二抗;采用Pierce ECL化学发光底物试剂盒,以胶片显影、定影。

表1 miR-150、IRAK2 mRNA检测中RT-PCR引物序列Table 1 RT-PCR primer sequences in miR-150 and IRAK2 mRNA detection
1.8 统计学处理
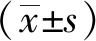
2 结果
2.1 行为学症状评分
NC组偶有抓鼻,AR组抓鼻、喷嚏及流涕症状明显,得分均>5分,表明建模成功,AR组症状评分(6.83±0.26)分高于NC组(2.36±0.31)分,差异有统计学意义(P<0.01)。
2.2 组织病理学改变
小鼠鼻腔病理组织切片,HE染色结果显示,NC组的鼻黏膜组织中,上皮细胞整齐排列有序,腺体组织结构正常,清晰可见, 腺体细胞形态正常,黏膜内小血管管壁完整清晰,周边未见炎性细胞渗出;AR组鼻黏膜纤毛大量脱落,组织间质水肿、小血管扩张、腺体增生,固有层内可见大量的炎性细胞浸润(图1)。
2.3 血清OVA-sIgE、IL-4、IL-13水平
ELISA测定AR组血清OVA-sIgE、IL-4水平均明显高于对照组,差异有统计学意义(P<0.001); AR组血清IL-13表达水平明显高于NC组,差异有统计学意义(P<0.01)(图2)。
2.4 两组小鼠miR-150相对表达量与IRAK2、OVA-sIgE、IL-4、IL-13含量的相关性分析
AR组和NC组miR-150相对表达水平与IRAK2、OVA-sIgE、IL-4、IL-13含量均成负相关(r=-0.841、-0.869、-0.834、-0.857,P<0.05)。
2.5 miR-150的结合位点
Target Scan Mouse6.2生物信息学软件预测IRAK2 mRNA是miR-150的靶基因,miR-150可与IRAK2 3′-UTR端特异性结合(图3)。
2.6 两组小鼠鼻黏膜组织miR-150和IRAK2 mRNA表达
qRT-PCR检测结果显示,与NC组相比,AR组miR-150表达下降,IRAK2 mRNA表达增高,差异有统计学意义(P<0.05)(图4)。
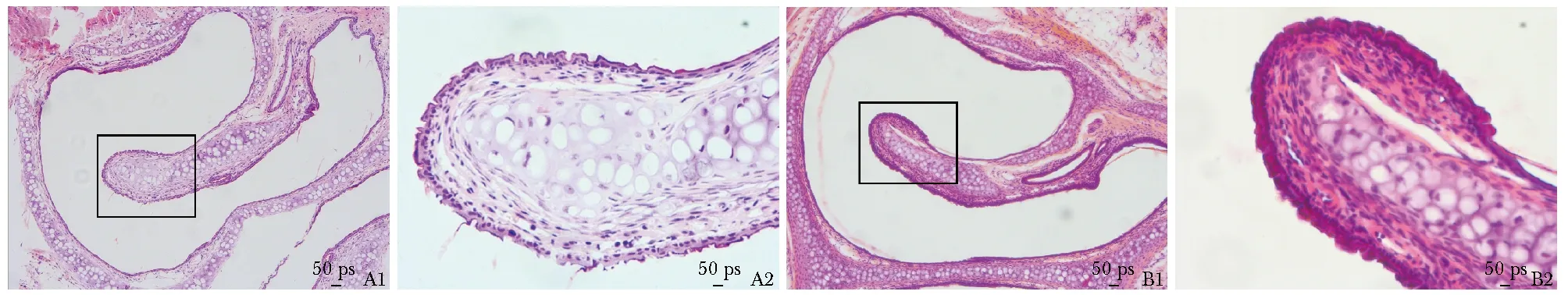
图1小鼠鼻腔组织HE染色(A1,B1 ×100;A2,B2 ×400)
Fig1HE staining of nasal tissues of mice (A1,B1 ×100;A2,B2 ×400)
A1,A2:正常对照组(NC);B1,B2:模型组(AR)

图2小鼠血清细胞因子表达水平
Fig2Expression level of cytokines in serum of mice
A:OVA-sIgE; B:IL-4; C:IL-13;与正常对照组比较,**P<0.001,*P<0.01
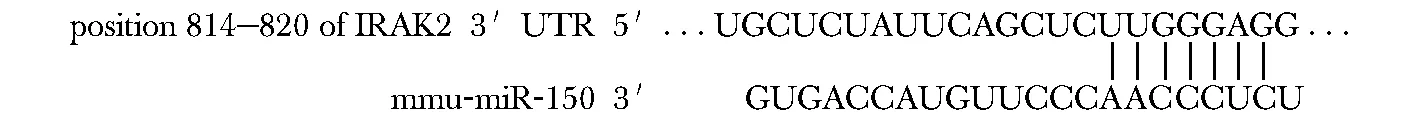
图3IRAK2 3′-UTR端包含一段miR-150基因预测序列
Fig3The end of IRAK2 3′-utr contains a predicted sequence of miR-150 gene
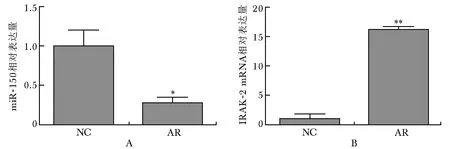
图4小鼠鼻黏膜miR-150和IRAK2 mRNA的表达水平
Fig4mRNA expression levels of miR-150 and IRAK2 in nasal mucosa of mice
A:miR-150 mRNA表达水平;B:IRAK2 mRNA表达水平; 与正常对照组比较,***P<0.001,**P<0.01,*P<0.05
2.7 miR-150表达与IRAK2表达的关系
Pearson相关分析表明,miR-150相对表达水平与IRAK2 mRNA的表达呈负相关(r=-0.841,P<0.05)(图5)。
2.8 两组小鼠鼻黏膜组织中IRAK2蛋白表达量
Western blot结果表明,与NC组相比,AR组小鼠鼻黏膜IRAK2蛋白表达增强,差异有统计学意义(P<0.05)(图6)。
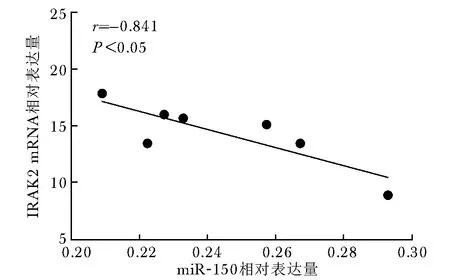
图5 miR-150相对表达量与IRAK2 mRNA相对表达量的相关性Fig 5 Correlation between miR-150 relative expression and IRAK2 mRNA expression
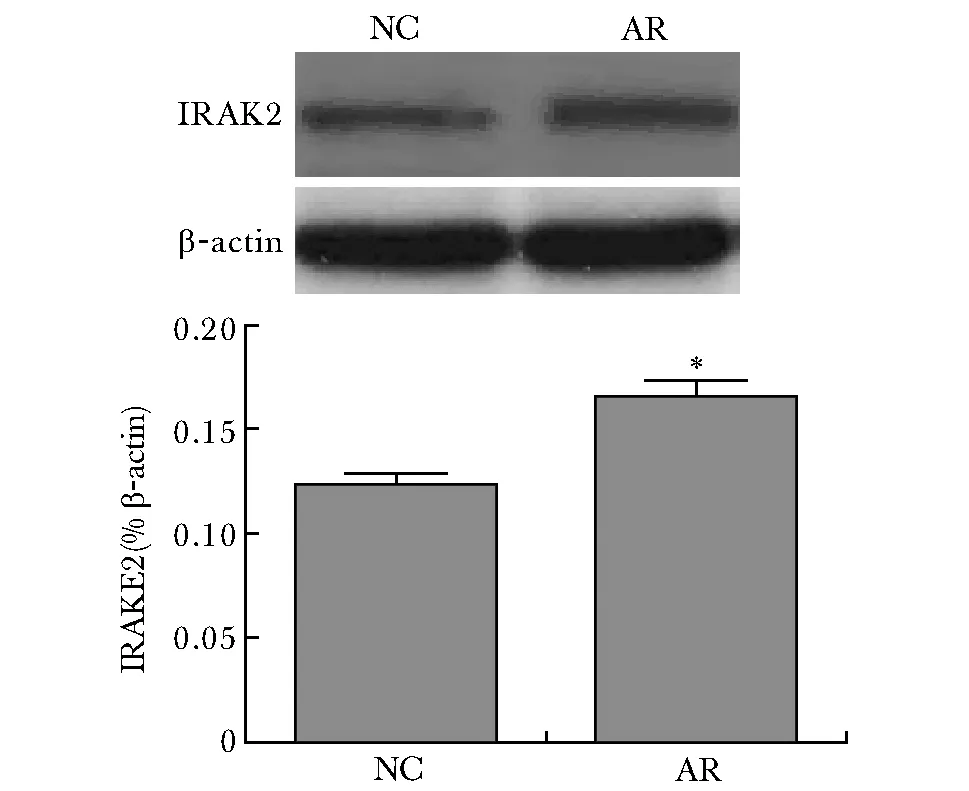
图6 小鼠鼻黏膜组织IRAK2蛋白表达水平Fig 6 Expression level of IRAK2 protein in nasal mucosa of mice
3 讨论
变应性鼻炎是耳鼻咽喉科的常见病,多发病,但其发病机制尚不完全清楚,目前以Finotto等[8]的Th1/Th2失衡理论被广为接受。Th2细胞优势分化和Th2细胞因子如IL-4、IL-5等过度表达是过敏性疾病的根本免疫学特征。miRNAs对过敏性疾病Th1/Th2平衡和细胞因子的影响已有报道[9],其在调节免疫进程中的作用越来越受到关注,从病理机制上提示了miRNA的重要调节作用,但特异性的 miRNA在变应性鼻炎中的作用及机制尚不清楚。
miR-150是一类由内源基因编码的单链小分子RNA,通过转录后调控抑制其靶基因蛋白的表达,在肿瘤侵袭与转移[10]、炎症反应[11-12]、抑制效应T细胞的生成[13]等多方面发挥着重要的调控作用。研究发现miR-150在静止成熟淋巴细胞(B细胞和T细胞)中高表达,其表达在随后分化为效应细胞Th1和Th2亚群时下降[14]。本研究结果表明miR-150在变应性鼻炎小鼠鼻黏膜中表达降低,这可能与变应性鼻炎发病过程中Th2细胞优势分化有关。此外,miR-150在炎症中的作用也引起了广泛关注,在脓毒症患者外周血白细胞中miR-150表达降低,且miR-150 含量越低,炎症反应越严重,预后越差[11]。进一步研究发现,miR-150通过抑制其靶基因Ang2的表达,降低脓毒症导致的血管损伤及其死亡率[12]。也有研究证明在革兰阴性细菌性败血症患者中发现miR-150下降,miR-150 可介导炎症反应过程,一定程度上可用于判断疾病严重程度及预后[15]。Nayoung等[16]研究表明,miR-150是抑制NK细胞裂解活性的一种常见的转录后调节因子,对小鼠和人NK细胞的穿孔素-1(perforin-1,PRF-1)有抑制作用。实际上,已经报道NK细胞参与了过敏性疾病[17-19],NK细胞可能有助于PRF-1介导的过敏反应[20]。除了PRF-1外,miR-150通过特异性结合多个下游靶基因参与多种信号通路的调控,包括c-Myb[21-22],EGR2[23]和FOXP1[24]等。本研究通过Target Scan等生物信息学检测,发现IRAK2 3′-UTR端包含一段miR-150基因预测序列,说明miR-150能够与IRAK2 mRNA特异性结合,降解IRAK2 mRNA,并抑制其翻译成为蛋白。进一步研究结果表明与正常对照组相比,AR组小鼠鼻黏膜miR-150表达降低,同时IRAK2 mRNA和蛋白表达水平增高,提示miR-150与其靶基因IRAK2之间可能存在某种相互作用,参与了变应性鼻炎的发生发展。
白细胞介素-1相关激酶(interleukin-1 receptor associated kinase,IRAKs) 家族是TLR介导信号通路中的重要蛋白激酶和调节分子,参与细胞内的信号网络控制和免疫反应,对于炎症反应、抗病毒作用、激活天然免疫应答及由此导致的自身免疫病性疾病和炎症性疾病的发生都有重要的调节作用,人类的各种炎症性疾病及自身免疫性疾病亦跟IRAKs基因的突变有关。目前研究表明IRAK2能够激活TLR2,TLR4和TLR9介导的信号通路,在维持TLR介导的NF-κB转录中起重要调节作用[25]。本研究显示miR-150的相对表达量与IRAK2 mRNA表达量和OVA特异性IgE、IL-4、IL-13的含量呈负相关,进一步说明miR-150与其靶基因IRAK2之间的相互作用与IL-4、IL-13等细胞因子异常密切相关,其机制可能是miR-150参与调控TLR介导的信号传导通路,影响相关细胞因子的表达。但由于本研究没有通过体内、外的miR靶基因实验(如过表达或敲减miR),后续仍需要进一步深入研究,才能明确miR-150与IRAK2之间的相互作用关系,及其调控有关信号通路的作用机制。
综上所述,miR-150能特异性结合其靶基因IRAK2,通过转录后调控抑制IRAK2的表达。变应性鼻炎miR-150表达下降与IRAK2表达增高,可能与变应性鼻炎的发生和发展有关,这对变应性鼻炎的个性化诊疗与靶向药物研发,具有良好的前景及潜在价值。
猜你喜欢
现代仪器与医疗(2022年4期)2022-10-08
中华实用诊断与治疗杂志(2022年1期)2022-08-31
健康体检与管理(2022年4期)2022-05-13
现代临床医学(2022年2期)2022-04-19
现代临床医学(2021年5期)2021-11-02
中老年保健(2021年2期)2021-08-22
中老年保健(2021年3期)2021-08-22
云南医药(2021年3期)2021-07-21
中国生殖健康(2020年2期)2021-01-18
中华养生保健(2020年2期)2020-11-16
