
定虫隆对锈赤扁谷盗生长发育影响及其作用机理初探
2020-03-09 03:21鲁玉杰杜梦园王争艳
环境昆虫学报 2020年1期
鲁玉杰,杜梦园,张 蒙,王争艳
((河南工业大学粮油食品学院,河南粮食作物协同创新中心,粮食储藏安全河南省协同创新中心,郑州 450001))
锈赤扁谷盗Cryptolestesferrugineus(Stephens)是当今世界上最难杀死的储粮害虫之一,它能够引起粮食局部发热霉变(苏青峰等,2013)。由于其长期单一使用磷化氢,导致锈赤扁谷盗对磷化氢的抗性急剧提高(曹阳,2006),目前磷化氢熏蒸已经很难彻底杀死锈赤扁谷盗。寻找新型的、对环境友好型的杀虫剂,已成当务之急。昆虫生长调节剂杀虫具有不污染环境、对人畜安全,以及对多数非靶标生物无害的优点(杨惠,2001),符合新型杀虫剂的要求。昆虫生长调节剂的种类很多,其中苯甲酰基脲类几丁质合成抑制剂以其独特的作用机制,对非靶标生物具有较高的选择性、使用浓度低、降解速度快、对环境友好等传统农业无法比拟的优点,被列为特异性的昆虫生长调节剂,已成为新农药创制的一个热门研究领域(罗会等,2009)。
定虫隆(chlorfluazuron)是一种苯甲酰基脲类几丁质合成抑制剂(冒德寿等,2001),能抑制害虫的几丁质合成,阻断蜕皮过程,致使蜕皮困难、取食量下降(梁英,2009),从而导致其死亡或不育(李秀峰和李玉新,2001;罗会等,2009)。因其独特的杀虫机制、较高的特异性、低残留等优点,被称为新型杀虫剂,常用来防治鳞翅目、鞘翅目等害虫(米娜等,2009)。研究表明,定虫隆能有效的抑制谷蠹Rhizoperthadominica、米象Sitophilusoryzae、锯谷盗Oryzaephilussurinamensis和赤拟谷盗Triboliumcastaneum4种储粮害虫的繁殖,具有很好的防治效果(Elek and Logstaff,1994)。相关的研究表明,除虫脲能有效的抑制蚊子Culicidae几丁质的合成,使幼虫不能形成新表皮,致使幼虫畸形而死亡(Cetinetal.,2006)。氟虫脲对东亚飞蝗Locustamigratoria的几丁质合成也有明显的抑制作用(王贵强等,1996),从而起到了防治害虫的效果。苯甲酰基脲类几丁质合成抑制剂的作用机理可能是特异性的干扰昆虫表皮几丁质的合成,但其作用于几丁质合成中的哪一环节至今尚不清楚。已有研究表明,几丁质合成抑制剂影响几丁质合成酶(Postetal.,1974),也有研究表明定虫隆并不直接抑制几丁质合成酶(Gangishettietal.,2009)。关于定虫隆的具体作用靶标及毒性机理,目前还没有明确的结论。
定虫隆作为一种几丁质合成抑制剂被广泛应用,而目前对于定虫隆的研究一般集中在杀虫活性及农药残留方面(朱烈等,2016),定虫隆对锈赤扁谷盗产生的影响,及可能的作用靶标尚没有系统性的研究。本研究以锈赤扁谷盗为对象,通过亚致死剂量的定虫隆处理锈赤扁谷盗,研究定虫隆对其生长发育的影响,及可能作用的靶标,为以后定虫隆用于储粮害虫的防治提供理论基础。
1 材料与方法
1.1 材料
1.1.1供试昆虫
锈赤扁谷盗试虫来源于山西吕梁国家粮食储备库,在河南工业大学储粮生态研究室的人工气候室培养多代,饲养温度30℃±1℃,相对湿度60%±5%,选择同一时间孵化的幼虫,转移至玻璃瓶中,饲喂人工饲料,饲料为全麦粉 ∶燕麦 ∶酵母粉的比例为5 ∶4 ∶1。
1.1.2主要试剂与仪器
定虫隆:含量99%,CSSBIO(中国)提供;RNA提取试剂盒Total RNA Miniprep Kit购自Axygen公司;荧光定量PCR试剂盒SYBR Premix Ex Taq购于TaKaRa公司;荧光定量PCR仪Step One Plus Real Time PCR(Thermo Fisher Scientific美国)。PCR引物合成由苏州金唯智生物科技公司完成。
1.2 定虫隆对锈赤扁谷盗的毒力测定
取1 mg定虫隆粉剂,用1 mL丙酮溶解,配制1 mg/mL定虫隆母液。在洁净的培养皿中,每10 g全麦粉(含5%酵母粉)中加入10 mL含有所需量定虫隆的丙酮溶液,将饲料配制成0.0050 mg/kg、0.0091 mg/kg、0.0166 mg/kg、0.0302 mg/kg、0.0549 mg/kg、0.1000 mg/kg 6个浓度梯度,并将悬浮液搅拌摇匀,然后将溶液放在通风橱下,直至丙酮完全挥发。使用和上述完全一样的饲料,只添加丙酮作为对照。每组放入30头锈赤扁谷盗2龄幼虫,每个浓度3个重复。每天记录幼虫死亡数并观察表型变化。
1.3 亚致死剂量下定虫隆对锈赤扁谷盗生长发育的影响
根据定虫隆对锈赤扁谷盗的毒力方程计算出锈赤扁谷盗的亚致死剂量LD10、LD20、LD30。用同样的饲喂方法,分别用LD10、LD20、LD303个浓度的定虫隆处理锈赤扁谷盗2龄幼虫,以丙酮作为对照,观察LD10、LD20和LD30下,定虫隆对锈赤扁谷盗幼虫发育历期、体重、几丁质含量的影响。几丁质含量测定采用氨基葡萄糖法(Lehmann and White,1975)。
1.4 定虫隆对几丁质合成酶表达量的影响
根据本实验室得到的锈赤扁谷盗几丁质合成酶1基因(CfCHS1)的转录组数据,和已经提交到GenBank的锈赤扁谷盗几丁质合成酶2基因(CfCHS2)的全长序列(GenBank登录号:MH234580),设计荧光定量PCR引物(表1)。以同样的饲喂方法,用LD30剂量的定虫隆处理锈赤扁谷盗3龄幼虫,每组30头虫,3组重复,以丙酮处理为对照。提取定虫隆LD30处理48 h后的锈赤扁谷盗3龄幼虫总RNA,反转录成cDNA,进行荧光定量PCR。qPCR反应体系:cDNA模板2 μL(<100 ng),SYBR Premix Ex Taq Ⅱ(2×)10 μL,PCR上引物(10 μM)0.8 μL,PCR下引物(10 μM)0.8μL,ROX Reference Dye(50×)0.4 μL,ddH2O 6 μL,总反应体系为20 μL。反应程序:95℃预变性30 s,95℃变性5 s,60℃退火延伸30 s,72℃,30 s,40个循环。每组样品进行3个重复,以目的基因在对照组中的表达量为1,以ARF1为内参基因。

表1 本文中所用引物的名称、序列
1.5 数据统计与分析
死亡率(%)=(死亡数/总虫数)×100
校正死亡率(%)=[(处理组死亡率-对照组死亡率)/对照组存活率]×100
统计分析采用SPSS软件进行统计分析和检验,P=0.05。
qPCR结果采用比较CT值得相对表达量法(2-ΔΔCt)计算基因的相对表达量。实验结果以平均数±标准差(SD)表示。表达量的差异显著性比较采用SPSS软件中的单因素方差分析(ANOVA,Tukey氏多重比较法,P<0.05)进行。
2 结果与分析
2.1 定虫隆对锈赤扁谷盗的杀虫活性
根据生物测定结果,将其与剂量对数值进行回归分析,建立定虫隆毒力方程:y=7.514+1.81x,r=0.974,计算得LD50为0.041 mg/kg,95%置信区间为0.025~0.059 mg/kg;LD95为0.331 mg/kg,95%置信区间为0.270~5.832 mg/kg。亚致死剂量LD10、LD20、LD30分别为0.008 mg/kg、0.014 mg/kg、0.021 mg/kg。
2.2 亚致死剂量定虫隆对锈赤扁谷盗生长发育的影响
经过亚致死剂量定虫隆处理的锈赤扁谷盗幼虫,各个发育历期以及蛹期的天数都是有所增加的。随着定虫隆剂量的增加,锈赤扁谷盗的发育历期明显延长(表2)。定虫隆剂量越高,处理时间越长,锈赤扁谷盗体重下降越显著;经定虫隆LD30长期处理的锈赤扁谷盗,体重可下降25%(表3)。低剂量的定虫隆就可以导致锈赤扁谷盗几丁质含量显著下降(P<0.05)(图1)。
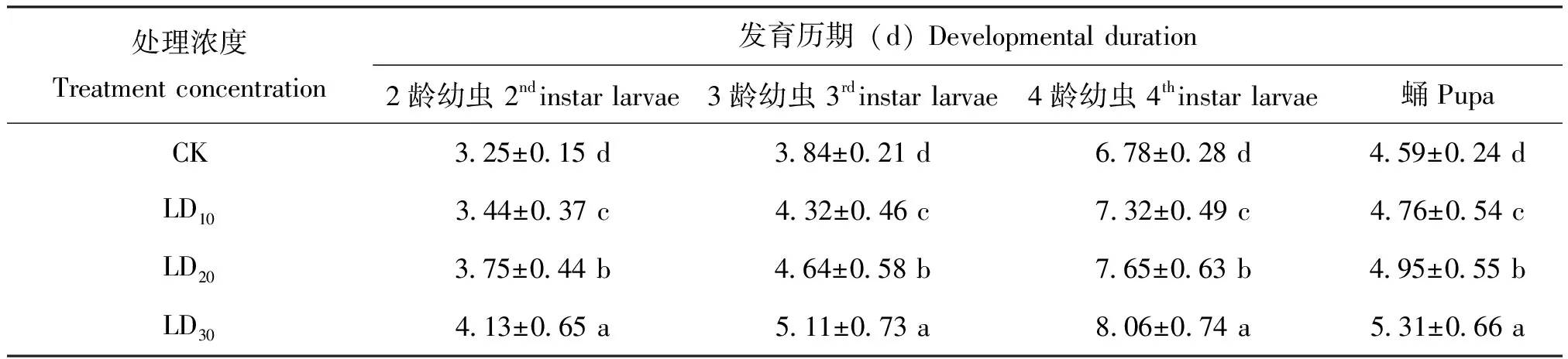
表2 不同浓度定虫隆处理后锈赤扁谷盗各虫态的发育历期
注:同列中不同的字母代表发育历期差异显著(P<0.05,Tukey’s B;n=3)。
Note: Different letters in the same column represent significant differences in development duration (P<0.05, Tukey’s B; n=3).

表3 不同浓度定虫隆处理后锈赤扁谷盗各虫态的体重
注:同列中不同的字母代表体重差异显著(P<0.05,Tukey’s B;n=3)。
Note: Different letters in the same column represented significant differences in body weight (P<0.05, Tukey’s B; n=3).

图1 定虫隆处理后锈赤扁谷盗各虫态几丁质的含量Fig.1 Content of chitin in different stages of Cryptolestes ferrugineus after treatment with chlorfluazuron注:柱上标的不同字母代表几丁质含量差异显著(P < 0.05,Tukey’s B;n=3)。Note: Different letters of the superscript on the column represent significant differences in the content of chitin (P < 0.05, Tukey’s B; n=3).
2.3 定虫隆处理后锈赤扁谷盗几丁质合成酶基因表达量的变化
用LD30定虫隆处理锈赤扁谷盗3龄幼虫,几丁质合成酶基因表达量的变化如图2所示。从图2可以看出,CfCHS1基因的表达量明显上调,是对照组的1.787倍。CfCHS2基因的表达量也明显上调,是对照组表达量的1.274倍。说明了定虫隆可以显著提高锈赤扁谷盗几丁质合成酶CfCHS1和CfCHS2基因的表达量。

图2 定虫隆处理后锈赤扁谷盗3龄幼虫几丁质合成酶CfCHS1和CfCHS2基因的表达量Fig.2 Expression level of chitin synthase CfCHS1 and CfCHS2 in the third instar larvae of Cryptolestes ferrugineus treatment with chlorfluazuron注:CK代表对照组,LC30代表处理组。数据均代表3次生物学重复的平均值±标准差。柱上标的不同字母代表基因表达差异显著(P<0.05,Tukey’s B;n=3)。Note: The data are means±SD.Histograms with different letters indicate statistically significant difference (P < 0.05, Tukey’s B; n=3).
3 讨论与结论
目前关于苯甲酰基脲抑制剂的亚致死效应研究相对较少,其作用机制也尚不清楚。本研究选取定虫隆进行试验,使用亚致死剂量持续处理锈赤扁谷盗幼虫,发现其各个虫态的发育历期都明显延长,体重下降,几丁质含量减少,并出现畸形死亡个体,这一结果与除虫脲对中华稻蝗Oxyachinensis的影响(曾慧花等,2008)相同,与灭幼脲能显著降低家蝇Muscadomestica几丁质的含量(Ishaaya and Casida,1974)的结果也是一致的。这说明定虫隆作为几丁质合成抑制剂,的确抑制了锈赤扁谷盗幼虫几丁质的合成,但发育历期的延长,和体重的下降,暗示着定虫隆可能对幼虫产生其他影响。
在灭幼脲处理家蚕Bombyxmori和四斑按蚊Anophelesquadrimaculatus中,其幼虫的体重均显著减少(Leonardietal.,1996;Zhuetal.,2006),这与本研究的结果相同。在除虫脲对中华稻蝗的研究中发现,除虫脲不仅抑制了中华稻蝗的体重的增长,还对其进食产生了抑制(曾惠花等,2008)。Clarke用除虫脲处理东亚飞蝗,发现除了表皮结构外,东亚飞蝗围食膜中的几丁质合成也受到了抑制(Clarkeetal.,1977)。根据前人的研究,几丁质合成酶2参与中肠围食膜的形成(Merzendorfer,2006)。刘晓健利用RNAi干扰技术,对飞蝗几丁质合成酶2基因进行沉默,其试虫的取食量也明显减少,体重降低(刘晓健等,2014)。根据大量研究推测,定虫隆可能对锈赤扁谷盗几丁质合成酶2产生影响,使其不能正常反应,致使围食膜的形成受到影响,从而产生了与RNAi干扰相似的结果。
为了探究定虫隆作用机理,本文在用定虫隆处理锈赤扁谷盗之后,利用qPCR技术,测定了几丁质合成酶基因相对表达量的变化。结果显示,亚致死剂量定虫隆处理后,锈赤扁谷盗几丁质合成酶1和酶2的表达量显著提高。为什么定虫隆可以使几丁质合成酶基因的表达量提高?我们推测,定虫隆可能作用于锈赤扁谷盗几丁质合成酶。定虫隆抑制了几丁质合成酶,使其不能正常的进行几丁质的合成,从而引起几丁质合成受阻,锈赤扁谷盗为了维持机体的稳定,提高几丁质合成酶基因的表达量,从而形成一种代偿性增加的反馈机制。Zhang and Zhu(2006)使用除虫脲处理四斑按蚊,发现其AqCHS1的表达量增加也显著提高。在氟虫脲对东亚飞蝗和中华稻蝗处理后,其CHS1基因的表达量都有显著的升高(刘晓健等,2010)。至于定虫隆是如何作用于几丁质合成酶,几丁质含量的减少与几丁质合成酶表达量增加之间的关系,需要我们进一步去研究。
本研究用定虫隆处理锈赤扁谷盗,结果表明定虫隆对锈赤扁谷盗的生长发育有明显的抑制作用,可以有效控制锈赤扁谷盗,至于如何在储粮环境中使用定虫隆,还有待进一步的研究,并需符合农药管理相关要求。定虫隆在抑制几丁质合成的同时,也使锈赤扁谷盗的几丁质合成酶基因表达量提高,这表明定虫隆可能作用于几丁质合成途径中的几丁质合成酶。这为以后利用定虫隆防治锈赤扁谷盗提供理论基础。
猜你喜欢
中国生物防治学报(2022年4期)2022-09-16
湖北农业科学(2021年22期)2021-12-13
科技视界(2021年32期)2021-12-08
广东农业科学(2021年6期)2021-07-27
信阳农林学院学报(2021年1期)2021-04-01
林业科技(2020年3期)2021-01-21
湖北农业科学(2020年17期)2020-11-09
农家科技下旬刊(2020年5期)2020-06-19
水产科学(2019年5期)2019-09-18
农村农业农民·B版(2016年7期)2016-10-21
