
昆虫几丁质酶及其应用研究综述
2022-09-16 05:56李春春苟玉萍张克信张强艳刘长仲
中国生物防治学报 2022年4期
李春春,苟玉萍,张克信,张强艳,刘长仲
(甘肃农业大学植物保护学院/甘肃省农作物病虫害生物防治工程实验室,兰州 730070)
关 键 字:几丁质酶;功能分析;害虫防治;应用
我国地域辽阔,自然环境复杂多样,有害生物种类繁多,数量巨大。其中害虫作为主要的有害生物之一,每年对我国农林牧草造成了巨大的经济损失[1]。长期以来,害虫的防治主要依赖于化学药剂,且药剂施用量大、施用次数多,以致害虫容易产生抗药性,导致防治困难,从而造成恶性循环。我国是农药使用量最多的国家之一,农药污染,有悖于生态环境保护、且对人畜产生残留毒害,农药减量增效是我国今后较长时期的一项艰巨任务[2]。因此,探索农林牧草害虫绿色防控的新途径尤为重要。
几丁质(Chitin),是由氨N-乙酰基葡萄糖(GlcNAc)通过β-1,4糖苷键连接而成的线性多糖聚合物,也称为壳多糖、甲壳素、甲壳质,不溶于水和多数有机溶剂,是自然界中贮量仅次于纤维素的一种含氮的多糖[3]。昆虫几丁质主要存在于昆虫的外骨骼、肠道、气管以及口器和唾液腺中,是昆虫体壁、气管和中肠围食膜等结构的重要组成部分,起着维持昆虫特定形态,抵御机械损伤和病原物侵害的重要作用[4,5]。几丁质的合成与降解在昆虫体内维持着动态平衡,如果几丁质的含量在昆虫某一时期升高或降低,则会影响昆虫的正常生长发育和形态构造。可见,几丁质对昆虫的生命活动而言是不可或缺的。然而,高等植物和动物并不具有几丁质代谢系统,因此,选择调控昆虫几丁质酶来破坏昆虫几丁质结构或几丁质代谢来防治害虫,具有很大的发展潜力。通过对昆虫几丁质酶相关基因的功能研究发现,沉默昆虫几丁质酶基因能打破昆虫几丁质代谢平衡,抑制几丁质的合成并导致昆虫蜕皮困难、出现高的死亡率[6-8],这表明昆虫几丁质酶能够作为害虫的控制靶标。因此,在害虫防治体系中,掌握昆虫几丁质酶调控机理对于制定新型防治策略与技术研发具有重要意义。
1 昆虫几丁质代谢途径
昆虫会定期蜕皮,脱落旧的外骨骼,并合成新的外骨骼,以便维持正常的生长发育。这种不断替换的过程就是由几丁质合成和降解酶系协同作用完成的昆虫几丁质代谢过程。昆虫体内几丁质生物合成途径以昆虫血糖——海藻糖为前提物,经过一系列的生理和生化反应,终由几丁质合酶(chitin synthase,CHS)合成几丁质。昆虫几丁质降解途径主要由几丁质酶将几丁质最终降解为N-乙酰氨基葡萄糖单体(GlcNAc),这些游离的GlcNAc能再次用于几丁质的重建(图1)[9,10]。

图1 昆虫几丁质的代谢途径[9,10]Fig. 1 Chitin metabolic pathway in insects[9,10]
2 昆虫几丁质酶
2.1 昆虫几丁质酶的基因克隆
昆虫几丁质酶(Chitinase,Cht)是一类具有抑制或降解几丁质潜力的水解酶,包括内切几丁质酶和外切几丁质酶2种[11]。广义上的昆虫几丁质酶是指内切几丁质酶,作用于β-1,4-糖苷键,把几丁质水解为可溶性的小分子寡糖和低聚糖,然后再由外切酶水解产生N-乙酰葡萄糖胺单体,用以形成新的表皮[12,13]。
1993年,Kramer等[14]首次从烟草天蛾Manduca sexta中克隆到一条长为2452 bp编码554个氨基酸的几丁质酶基因。近年来,随着测序技术飞速发展,越来越多昆虫几丁质酶基因被克隆和验证,如黑腹果蝇Drosophila melanogaster、赤拟谷盗Tribolium castaneum、褐飞虱Nilaparvata lugens等,且不同昆虫拥有的Cht基因数量有所不同(表1)。
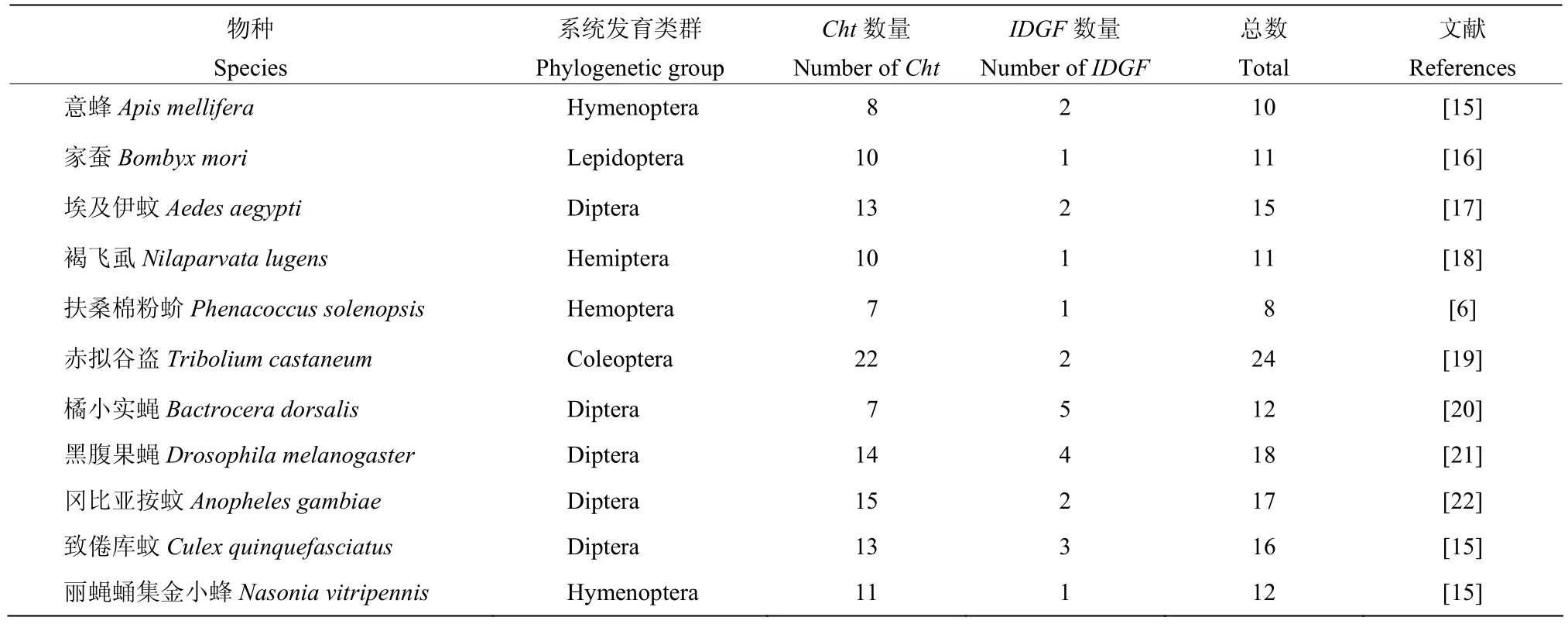
表1 几种昆虫的几丁质酶基因(Cht)数量Table 1 Number of chitinase genes (Cht) in several insects
2.2 昆虫几丁质酶及其编码蛋白的结构与性质
昆虫几丁质酶的分子量在45~85 kDa,等电点在5.0~7.0,大多属于18家族几丁质酶[23]。昆虫几丁质酶和几丁质酶样蛋白存在四个保守结构域:信号肽(Signal peptide, SP)、催化区域(Catalytic domains,CD)、富含脯氨酸、谷氨酸、丝氨酸和苏氨酸的S/T连接区域(Serine/Threonine-Rich Linker Region,S/T Linker)和C端几丁质结合区域(Chitin-binding domain,CBD)[24],以模式昆虫果蝇为例,其几丁质酶样蛋白的结构域如图2所示。基于系统发育分析、时空表达特异性和蛋白结构域特点,昆虫Cht和Cht类似蛋白主要划分为八个组(Group I~Ⅷ),其各组的几丁质酶序列结构特征如表2所示[25,26]。
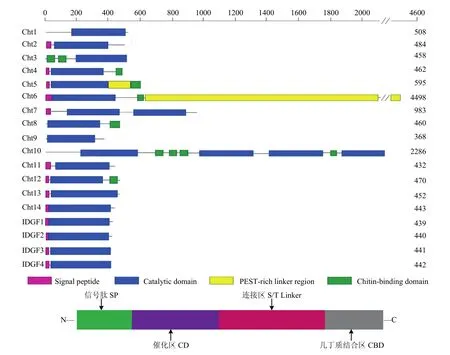
图2 果蝇几丁质酶样蛋白的结构域[21]Fig. 2 Domain architecture of identified Drosophila chitinase-like proteins[21]
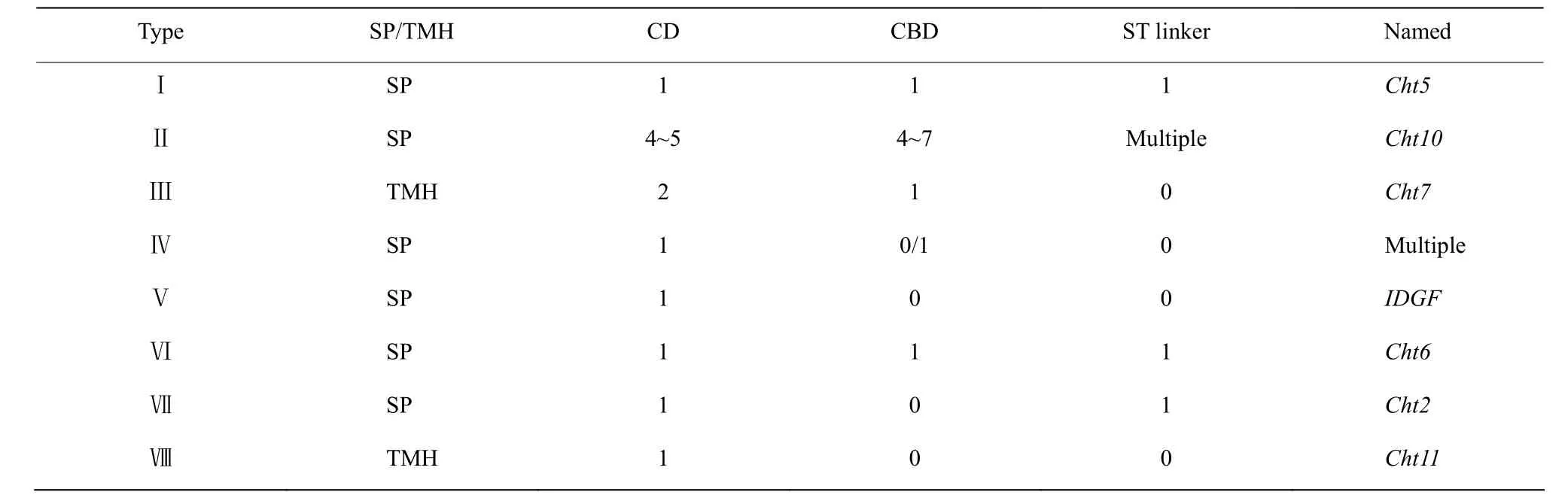
表2 昆虫几丁质酶I~VIII不同类型的序列结构域[25,26]Table 2 Domain architecture of group I—VIII insect chitinases[25,26]
3 昆虫几丁质酶基因调节昆虫几丁质代谢
3.1 昆虫几丁质酶调控机制的多样性与复杂性
昆虫几丁质酶基因的表达具有特异性。Cht5和Cht10基因在赤拟谷盗和冈比亚按蚊的肠道组织几乎不表达,但在体壁组织具有较高的mRNA水平[19,22];然而,云杉色卷蛾Choristoneura fumiferana的肠道组织中能检测到几丁质酶,但在表皮中检测不到与蜕皮相关的几丁质酶cDNA[27],说明肠道和表皮可能表达不同的组织特异性几丁质酶。MsCht1仅在烟草天蛾雄虫精巢和雌虫卵巢中特异性高表达[28],BdCht1在橘小实蝇卵巢组织也具有较高的表达量[29]。与此不同,在褐飞虱雌虫生殖器官高表达的几丁质酶基因主要为NlCht2和NlCht6[18],这说明物种差异使得在参与卵发育过程中的几丁质酶基因有所不同。此外,不同的几丁质酶基因在昆虫不同发育阶段的表达也存在特异性。扶桑棉粉蚧PsCht2在2龄若虫高表达,PsCht10、PsCht3-3和PsIDGF在雌性成虫中高表达,而PsCht4和PsCht4-1在雄性蛹和雄性成虫中高表达[6]。橘小实蝇BdCht1、BdCht7和BdCht10在幼虫生长和蜕皮过程中均有较高表达量[29]。
昆虫几丁质酶在昆虫蜕皮过程中发挥重要作用,但并非所有几丁质酶都在蜕皮的各个阶段发挥作用。在橘小实蝇中,在幼虫体壁组织高表达的基因有BdIDGF和BdCht2,而其他的几丁质酶基因均在成虫体壁组织具有较高的表达量,BdCht8在成虫中肠组织特异性高表达[29]。赤拟谷盗的TcCht10有助于幼虫-幼虫、幼虫-蛹和蛹-成虫阶段的蜕皮,而TcCht5只在蛹-成虫蜕皮中起作用[19],这表明几丁质酶调控机制存在多样性和复杂性。
3.2 昆虫几丁质酶基因的功能分析
几丁质酶是在昆虫蜕皮过程中用来消化其外骨骼和肠道内壁中的一组蛋白质[30],其功能涉及生长发育、蜕皮、食物消化、生殖和先天免疫等多个方面。它们能够与昆虫肠道和角质层内围食膜的几丁质成分结合,导致含有几丁质基质的降解和生物体的死亡[31,32]。目前,大量研究通过体外合成dsRNA,并经饲喂或注射方式使幼虫摄取,干扰相应几丁质酶基因的表达,来观察其对昆虫不同发育阶段的影响。
白背飞虱Sogatella furcifera[33]、黏虫Mythimna separata[34]等昆虫的Cht7基因在昆虫蜕皮前后表达量变化显著,说明Cht7基因与该昆虫的蜕皮相关;同样,当RNAi成功抑制黑腹果蝇DmCht7基因的表达后,该虫外形发生明显变化,1龄幼虫的表皮发生破裂,2龄和3龄幼虫的头壳和身体后部的气门发生了明显的变形,且在蛹-成虫的分化阶段易出现蜕皮困难、成虫不能成功展翅等现象[35]。赤拟谷盗和褐飞虱中的4~5个几丁质酶家族成员能够对其产生致死表型:在赤拟谷盗中,TcCht5在蛹-成虫期起重要作用,TcCht10与幼虫-幼虫、幼虫-蛹、蛹-成虫和卵的孵化有关,TcCht7在成虫翅原基及翅鞘的发育过程中发挥关键作用[19,36];在褐飞虱中,NlCht1、NlCht5、NlCht7、NlCht9、NlCht10能够影响若虫蜕皮,使褐飞虱出现不同程度的致死表型[37];RNAi褐飞虱的NlCht2、NlCht3、NlCht6、NlCht8和NlCht7后,该虫足前附肢的趾钩消失以致影响行走和弹跳,最终导致死亡[36,38]。同样,BdCht5能调控橘小实蝇幼虫-蛹和蛹-成虫变态过程中几丁质的降解[39];PsCht4或PsCht4-1能导致扶桑棉粉蚧蜕皮失败,沉默PsCht5导致化蛹缺陷,饲喂dsCht10导致3龄若虫体型变小,5 d后全部死亡[6]。最新研究发现,DmCht10可控制果蝇翅角质层中几丁质的数量和组织,抑制DmCht10后,果蝇翅表皮几丁质总量过度累积,进而导致几丁质的排布紊乱,片层结构破坏,不利于昆虫角质层和翅表皮的形成[40]。
综上所述,无论各个Cht具有怎样的差别,都能影响昆虫几丁质代谢,并且造成一定的昆虫死亡率。几丁质酶不仅有其特定的功能,而且在整个机体内相互作用,共同调控着昆虫几丁质的运行,通过昆虫几丁质酶调控昆虫生长发育来防治害虫,是一种潜在的生物杀虫途径和方法。
4 影响昆虫几丁质酶活性的因素
4.1 温度与pH
大多数昆虫几丁质酶的最适pH约为4~10,最适温度约为40 ℃~60 ℃,热稳定性一般在4 ℃~60 ℃[41]。然而,当作用底物不同时,几丁质酶活性也存在差异。例如,使用寡聚底物4MU(GlcNAc)3时,飞蝗LmCht5-1和LmCht5-2的最适pH为6;使用多聚底物CM-Chtitin-RBV时,二者的最适pH为9[42]。此外,温度对几丁质酶具有调控作用。对橘小实蝇的最新研究发现,在高温组(45 ℃处理1 h)中,成功存活昆虫的IDGF4表达量是对照组(25 ℃处理5 h)的3.15倍,用1000 ng/μL浓度的dsIDGF4处理橘小实蝇3龄幼虫48 h后,与dsGFP饲喂和水相比,IDGF4沉默率分别为69.2%和73.6%,成活率分别下降29.1%和27.1%,该基因表达模式的增加可能揭示了其参与热耐受性,高温可上调橘小实蝇IDGF4的基因表达,沉默该基因可降低橘小实蝇在高温胁迫下的存活率[43]。然而,小胸鳖甲Microdera punctipennis成虫在4 ℃和-4 ℃不同温度处理下发现,-4 ℃在诱导几丁质酶基因(MpCht4、MpCht16、MpCht17、MpCht5-B、MpCht13和MpIDGF2)表达方面比4 ℃具有更强的效果[44],表明昆虫几丁质酶可能具有多种功能,并在昆虫的冷适应中发挥作用,但其作用机制还有待进一步研究。
4.2 抑制剂
几丁质酶抑制剂能够特异性地结合并抑制几丁质酶的活性,使生物正常生长发育不能顺利进行,从而达到防治病虫害的效果[45]。目前已研究报道的几丁质酶抑制剂包括阿洛氨菌素(allosamidin)、精氨芬(argifin)、环二肽(CI-4)、阿尔加定(argadin)、甲基黄嘌呤(methvlxanthine)类及其衍生物等[46]。其中,阿洛氨菌素是从放线菌Streptomycessp.中分离获得的一种天然的假三糖,属糖基类化合物,由一个allosamizoline 基团和两个N-乙酰-D-allosamine基团组成,是最早最经典的Cht抑制剂,可有效地抑制不同来源的Cht,阻止昆虫几丁质酶的降解[47,48]。研究发现,抑制剂阿洛氨菌素对幕衣蛾Tineola biselliella幼虫的致死率很高[49],说明阿洛氨菌素可有效抑制幕衣蛾几丁质酶的降解,影响了该虫正常的生长发育。此外,阿洛氨菌素对斜纹夜蛾Spodoptera litura几丁质酶的抑制效果是家蚕的3~5倍[50]。这说明,阿洛氨菌素对不同昆虫几丁质酶的效果也有所差别。
4.3 蜕皮激素
昆虫20-羟基蜕皮酮(20-hydroxyecdysone,20E),是最活跃的蜕皮激素,其滴度在蜕皮前升高,触发与蜕皮相关基因的表达,控制和协调昆虫的发育。蜕皮激素的滴度影响昆虫体内几丁质代谢过程中多种酶的表达及活性。20E可诱导多种昆虫几丁质酶的表达。用20E处理黏虫幼虫后,不同时间点MsCht7基因表达量存在显著差异,MsCht7基因表达量在1~24 h随时间逐渐升高,24 h时最高,之后表达量迅速降低,观察其生物学发现,不同浓度20E处理后5龄幼虫蜕皮时间均提前,说明黏虫MsCht7基因表达量受蜕皮激素调控[34]。此外,家蚕5龄幼虫被注射4 µg 20E 6 h后,BmCht5基因表达量显著增加,并在8 h时达到最大值,而2 µg 20E诱导的BmCht5表达没有显著上调[51],蝗虫LmCht5基因[52]也能受到20E的调控。与之相类似,20E处理橘小实蝇幼虫12 h后,BdCht2、BdCht5、BdCht8、BdCht10和BdCht11的表达水平显著上调,BdCht7、BdIDGF1、BdIDGF2和BdIDGF3的表达量显著下调,表明参与幼虫生长发育的几丁质酶基因受蜕皮激素调控,至于是直接调控还是间接调控,尚需更多的研究结果回答[29]。
5 昆虫几丁质酶在害虫防治方面的应用前景
5.1 昆虫几丁质酶的生物杀虫剂
一些研究也表明,虽然昆虫几丁质酶能够调控昆虫体内几丁质的代谢平衡,但当其单独作用时杀虫效果不显著。如几丁质酶单独作用黏虫和甜菜夜蛾时,其杀虫活性不到10%,与空白组没有显著差异[53]。研究者认为,昆虫几丁质酶与其他杀虫剂协同作用时能拓宽杀虫谱,降低单虫毒素基因产生抗性的风险,显著增强杀虫效果,同时可以减少杀虫剂量,降低生态威胁。如几丁质酶基因Cht36与苏云金杆菌(Bacillus thuringiensis,Bt)杀虫剂协同对斜纹夜蛾幼虫的半致死浓度降低了30%[54]。此外,昆虫几丁质酶与杆状病毒协同作用,能够提高杆状病毒的杀虫能力。Shapiro等[55]研究发现昆虫几丁质酶能显著增强杆状病毒LdMNPV对舞毒蛾Lymantria dispar的致死作用,且暴露于NPV-几丁质酶组合的昆虫比仅摄入NPV的昆虫死亡得更快。同样,烟草天蛾几丁质酶的苜蓿银纹夜蛾核型多角体病毒(Autographa californica multiple nucleopolyhedrovirus,AcMNPV),通过在烟草天蛾幼虫中的过表达产生几丁质酶来提高杀伤速度[56]。以上研究结果表明,昆虫几丁质酶可以影响昆虫的生长发育以达到害虫防治目的,具有成为新型高效的生物杀虫剂的潜力,而且这种的生物杀虫剂对环境友好、人畜无害,是今后研究与开发安全、高效生物农药的热点领域。到目前为止,具有良好杀虫活性的基因还比较少。因此,几丁质酶也可以用于开发,从而减少合成农药对生态系统的引入和影响。
5.2 昆虫几丁质酶在转基因作物中的应用
昆虫几丁质酶可增强植物对害虫的抗性能力。研究发现,细菌和植物的几丁质酶基因导入转基因作物中可增强其对病原菌的防御能力,但对害虫抵御效果不显著,而将昆虫几丁质酶基因导入作物后,得到的转基因作物能高表达具有生物活性的几丁质酶,可以显著提高作物的防虫效果[23]。研究表明,表达昆虫几丁质酶基因的转基因植物对昆虫取食的抗性增强,这是因为它能够降解由β-1,4-n-乙酰氨基葡萄糖组成的几丁质线性聚合物,这是昆虫角质层和PM的重要组成部分。因此,表达昆虫几丁质酶的转基因作物可以免受病原真菌和害虫的侵害。如海灰翅夜蛾Spodoptera littoralis中几丁质酶基因的转基因玉米植株对玉米螟Pyraustanubilalis的抗性增加了约50%[57]。引入抗虫基因的转基因植物为害虫防治提供了一种有效的替代方法。Ding等[58]将产自烟草天蛾的几丁质酶基因引入烟草中,转基因烟草植株对绿棉铃虫Heliothis virescens幼虫侵染表现出较强的抗性。烟草天蛾几丁质酶基因的转基因番木瓜系在田间条件下对蜘蛛和螨也表现出显著的耐受能力[59]。
此外,研究者发现将昆虫几丁质酶基因与其他抗虫基因同时转入作物中时,杀虫活性与单一抗虫基因相比显著增强。通过农杆菌介导将含有昆虫特异性几丁质酶基因(Chi)和蝎子毒素基因(BmkIT)的新型抗虫基因组合导入甘蓝型油菜中,几丁质酶和蝎毒素蛋白均高表达,对供试害虫具有较强的抗性,当只有蝎子毒素基因表达量较高时,甘蓝型油菜则表现出中等杀虫活性[60]。Regev等[61]也证实了 Bt毒素与几丁质酶的协同作用可以增强对斜纹夜蛾幼虫侵染的抗性。因此,昆虫几丁质代谢关键酶的抗虫基因组合可作为农作物转基因的新基因来源之一,是害虫防控的新思路,有广阔的应用前景。
6 昆虫几丁质代谢在害虫防治中的不足与问题
6.1 昆虫几丁质提取困难
昆虫几丁质在农林[62]、医药[63]、食品[64]等方面具有巨大的应用价值和广泛的市场,但昆虫几丁质的提取面临着巨大挑战。目前,缺乏一套高效、环保、批量提取昆虫几丁质的工艺流程,因此,今后需要进行大量的科学研究完善昆虫几丁质的提取工艺。例如,在未经处理的幼虫,甚至包括漂白和干燥的幼虫,由于其体内脂肪含量较高(3%~20%)[65],几丁质通常不显示,这一阶段的昆虫细胞壁具有疏水排斥作用,使用磷酸提取几丁质的效果不显著。此外,昆虫细胞中的色素含量也影响几丁质的提取[66]。研究者发现,黑色素在蛹期或昆虫后期能与几丁质共价结合,影响使用有机酸(H3PO4)提取几丁质[67]。所以,开发新颖、高效的昆虫几丁质提取技术已迫在眉睫。
6.2 昆虫几丁质代谢应用的局限性
几丁质降解酶的转基因作物表达仅能将害虫中肠围食膜作为靶点,因此,创建靶向几丁质酶结构的RNAi-植物保护剂是害虫防治的新思路,此方法可将dsRNA传输到一些昆虫的整个虫体内,如鞘翅目,使得害虫整个虫体的几丁质结构成为靶标[68]。这些针对几丁质代谢途径的基于RNAi的研究大多数处于概念验证阶段,且并不是所有害虫和基因对dsRNA都会做出反应,因此还需要进一步研究克服不同害虫中RNAi效率的局限性,然后才能被广泛应用[69,70]。此外,昆虫几丁质酶虽已广泛应用于转基因作物,但在一些转基因植物中几丁质酶的转录水平低,易被降解,并对非靶标生物存在副作用等问题[71]。而且,除昆虫几丁质酶之外,其他昆虫几丁质代谢关键酶在转基因作物中的应用研究实例较少。但随着科学技术的不断发展,昆虫几丁质代谢的关键酶基因会更加广泛地应用在植物保护领域。
7 小结与展望
昆虫几丁质酶能够干扰昆虫的几丁质结构,消化昆虫营养基质和外骨骼中的几丁质,这对昆虫的生长、发育和生存至关重要,在害虫防治方面具有特别的效果。就目前研究而言,通过体外合成dsRNA,并经饲喂或注射方式使害虫幼虫摄取,破坏昆虫几丁质代谢平衡,干扰相应几丁质酶基因表达,导致害虫蜕皮困难、翅发育畸形,甚至死亡,是一种潜在的生物杀虫途径[72]。几丁质酶在害虫防治方面的应用前景主要体现在两方面,一是昆虫几丁质酶可作为生物杀虫剂备受人们的关注,当昆虫几丁质酶与其他杀虫剂协同作用时杀虫效果更加。然而,目前研究发现,几丁质酶调控害虫的机制复杂多样,且其活性受多种因素影响,因此实践研究尚未广泛开展,还需要进一步探索;二是昆虫几丁质酶转基因作物具有很好的抗虫效果,所以,昆虫几丁质代谢关键酶的转基因作物将是害虫防控的新思路,有较为广阔的应用前景。
综上所述,针对昆虫几丁质合成和降解酶开发的用于害虫防治药物,在植物保护领域具有潜在的应用价值。探索昆虫几丁质酶对昆虫的调控机制可能为设计既对环境无害又对害虫高效的新型杀虫剂提供机会,为害虫防治提供新的策略和思路[73]。
猜你喜欢
军事文摘(2022年12期)2022-11-03
天津农业科学(2022年6期)2022-07-17
今日农业(2022年3期)2022-06-05
发明与创新·小学生(2021年10期)2021-10-21
好孩子画报(2021年9期)2021-09-26
新少年(2020年8期)2020-10-29
农村农业农民·B版(2016年7期)2016-10-21
少年科学(2009年11期)2009-07-09
杂文选刊(2007年16期)2007-05-14
中国青年(1965年19期)1965-08-20
