
5型腺病毒E1A基因通过调控GP73的表达抑制肝癌细胞增殖的初步研究
2019-09-27 02:26李栋宇魏从文宫静钟辉汪浩勇
生物技术通讯 2019年4期
李栋宇,魏从文,宫静,钟辉,汪浩勇
1. 湖北工业大学,湖北 武汉430068;2. 军事医学研究院 生物工程研究所,北京100850;3. 安徽大学,安徽 合肥230000
人5 型腺病毒(adenovirus type 5,Ad5)早期区域1A(early region 1A,E1A)基因是近年新发现的一种抑癌基因,其通过2 个外显子的可变剪接来编码2 种主要蛋白质,而这2 种早期病毒蛋白(长度为243~289 个氨基酸残基)可以激活或抑制几种病毒或细胞基因的转录,从而调节细胞周期到达抑癌作用[1-2]。Ad5E1A 还具有转化和反式激活活性。通过直接竞争RB1 蛋白上的相同结合位点,诱导从RB1 中分离出E2F1 转录因子,E2F1 的释放导致ARF 蛋白介导的MDM2 被抑制并导致TP53/p53 积累,因为它不再是MDM2 介导的泛素化降解的目标。反过来,TP53 的增加将阻止细胞增殖并介导其死亡[3]。虽然Ad5E1A 已被确定是一种有效的转录激活因子,但人们对其如何改变细胞转录知之甚少[4]。原发性肝癌(肝癌)恶性程度高,预后极差,手术、化疗的疗效均不佳,近年来基因治疗和细胞治疗已成为有效的新型疗法。现有研究表明,Ad5 对肝癌细胞有较强的生长抑制作用[5],但是其抑制机制尚不明确。
高尔基蛋白73(Golgi protein 73,GP73)是一种跨膜糖蛋白,2000 年由Kladney 等[6]发现。研究发现,GP73 可作为肝癌诊断的标志物[7-8]。GP73在正常肝组织中几乎不表达,但在各种原因引发的肝脏疾病中,几乎所有肝细胞均有表达,特别是结缔组织周边和肝硬化结节部位表达尤其强烈[9]。70%以上的肝癌病人,GP73 蛋白在血清及肝组织中的表达水平均显著上调[10]。多篇文章报道GP73 在肝癌组织和血清中呈高表达,张宏冰等发现mTORC1 能够通过调节GP73 促进肝癌的发生发展[11]。已有研究证实了GP73 在肝癌发生发展及转移中的重要作用。我们前期研究发现,GP73 对肿瘤微环境具有重要的调节作用。之前研究未报道Ad5E1A 与GP73 的相互作用,因此探讨Ad5E1A 基因对GP73 的表达具有重要的科学意义。本研究中,我们在肝癌细胞HepG2 中过表达Ad5E1A 基因,并观察其对GP73 表达水平的影响以及肝癌细胞的增殖情况,为下一步研究肝癌发生机制,开发新型治疗方案提供实验基础。
1 材料与方法
1.1 材料
人胚胎肾细胞HEK293T、肝癌细胞HepG2(本实验室保存);pcDNA3-Flag-GP73(本实验室构建);腺病毒(李山虎教授赠与);大肠杆菌DH5α(北京擎科生物科技有限公司);限制性内切酶EcoRⅠ、BamHⅠ,T4DNA 连接酶(TaKaRa 公司);胶回收试剂盒(Promega 公司);质粒提取试剂盒(天根公司);DMEM 培养基(Gibco 生物公司);胎牛血清(杭州四季青公司);anti-Myc 抗体、anti-Flag 抗体、anti-Tubulin 抗体(Sigma-Aldrich 公司);脂质转染试剂jetPRIME(Polyplus 公司);100 mmol/L 青霉素(华北制药集团);100 mmol/L 链霉素(山东鲁抗集团);引物合成和测序由北京擎科生物科技有限公司完成。
1.2 Ad5E1A基因真核表达载体pcDNA3-Flag-Ad5E1A(Flag-Ad5E1A)的构建
以腺病毒为模板进行PCR 扩增,琼脂糖凝胶电泳鉴定并切胶回收PCR 产物。PCR 产物和pcDNA3-Flag-Vetor 质粒分别经EcoRⅠ、BamHⅠ双酶切,酶切产物于室温下连接后转化感受态大肠杆菌DH5α,在氨苄青霉素筛选压力下于LB 培养板37℃培养过夜,挑取单克隆在LB 培养基中扩增,进行菌液PCR 鉴定及测序。通过NCBI Blast进行序列比对,序列正确的质粒命名为pcDNA3-Flag-Ad5E1A。
1.3 细胞培养、转染及免疫印迹分析
细胞培养于含10%胎牛血清的DMEM 中,转染前24 h 细胞接种于培养皿中。转染前1 h 更换一半新鲜的DMEM 培养基,按说明书要求配制质粒和转染试剂混合液,滴加入细胞,常规培养,4~6 h 后补充另一半培养基。转染后24 h 收集细胞,1×PBS 冲洗3 次,3000 r/min 离心3 min 后弃上清。加人适量NP-40 裂解液[10 mmol/L Tris-HCl(pH8.0),1 mol/L NaCl,1 mmol/L EDTA,1%NP40],冰上裂解细胞15 min,12 000 r/min 冷冻离心后取上清,加入4×SDS 缓冲液[10%甘油,2%十二 烷基 磺 酸钠,50 mmol/L Tris-Cl(pH6.8),2.5% β-巯基乙醇,0.1%溴酚蓝],沸水浴10 min,12 000 r/min 离心10 min,取上清进行SDSPAGE,半干法转到PVDF 膜上,用5%脱脂牛奶液室温封闭1 h,一抗和二抗分别室温孵育1 h,TBST 洗3 次,每次5 min,将Perkin Elmer 的ECL发光液均匀涂在PVDF 膜上,暗室显影。
2 结果
2.1 pcDNA3-Flag-Ad5E1A真核表达载体的构建
按1.2 所述方法,首先对目的基因Ad5E1A 进行PCR 扩增,琼脂糖凝胶电泳鉴定结果见图1A,在750~1000 bp 处出现特异扩增条带,与目的片段Ad5E1A(870 bp)相符。构建pcDNA3-Flag-Ad5E1A 真核表达载体,随机挑选平板上的3 个单克隆菌落,分别编号为1、2、3,在含氨苄青霉素的LB 培养基中扩增后进行菌液PCR 鉴定,琼脂糖凝胶电泳结果见图1B,5-1、5-3 泳道750~1000 bp处出现特异性扩增条带,与目的片段Ad5E1A 相符。选取5-1、5-3 号对应的菌液测序,结果正确,命名为pcDNA3-Flag-Ad5E1A。
2.2 Flag-Ad5E1A在HEK293T细胞中的表达
将Flag-Ad5E1A 质粒转染HEK293T 细胞,通过免疫印迹检测重组质粒的表达,结果如图2,构建的真核表达质粒在HEK293T 细胞中均得到表达,产物相对分子质量约33×103,与预期相符。
2.3 Ad5E1A基因过表达后对HepG2细胞中GP73表达的影响
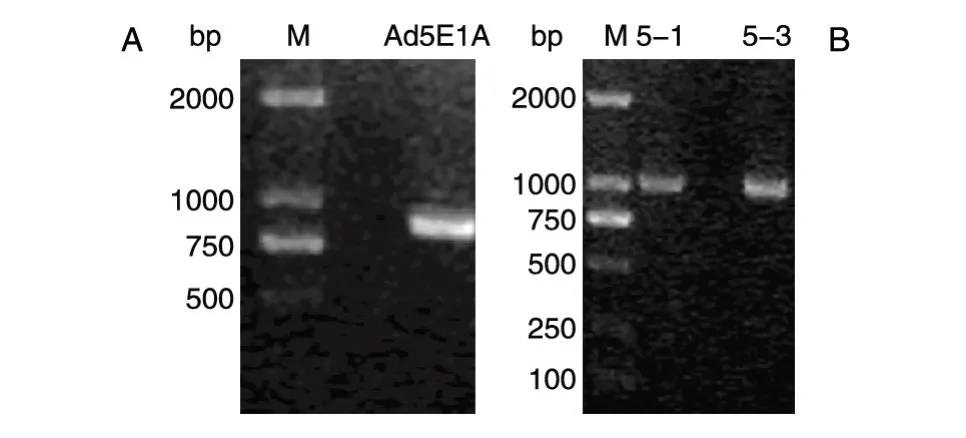
图1 琼脂糖凝胶电泳检测Ad5E1A 基因扩增产物(A),菌液PCR 鉴定插入目的基因片段(B)
实验组为过表达GP73 的HepG2 细胞中转染Ad5E1A 基 因,对 照组为 过表达GP73 的HepG2 细胞中转染Flag-Vector,各组均在37℃、5% CO2条件下培养,24 h 后收集细胞,用含10% β-巯基乙醇的SDS 裂解液裂解细胞,进行免疫印迹实验。结果见图3,与对照组相比,实验组的GP73 表达水平明显降低。
2.4 pcDNA3-Flag-Ad5E1A抑制GP73表达进而影响HepG2细胞增殖
将HepG2 细胞均分为4 组,用酶标仪测定D450nm值,分析细胞增殖情况。与转染Flag-GP73组相比,Ad5E1A 组的抑制率为17.5%,差异无统计学意义(P>0.05);而与转染Flag-GP73 组比,共转Ad5E1A 和GP73 组的抑制率为37.8%,差异有统计学意义(P<0.05)(图4)。因此可以看出,Ad5E1A 基因的抑制HepG2 细胞增殖作用与GP73相关。抑制率(%)=(对照组D450nm值-实验组D450nm值)/对照组D450nm值。
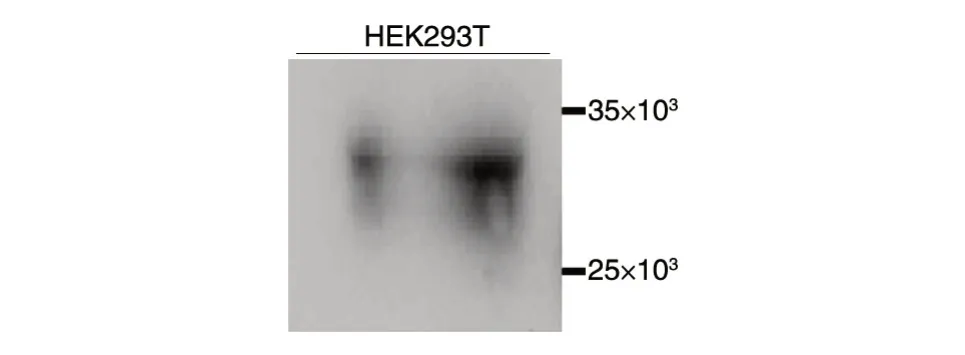
图2 HEK293T 细胞中Flag-Ad5E1A 蛋白的表达

图3 Ad5E1A 基因过表达对HepG2 细胞中GP73 表达的影响

图4 单独转染GP73 和共转GP73、Ad5E1A 时HepG2 细胞的增殖情况
3 讨论
肝癌作为发病率和病死率极高的癌症之一,一直都是科研人员以及临床工作者努力攻克的难题[12]。GP73 是维持动物正常生存以及肝肾生长发育的重要因子。近年来的研究发现GP73 有可能成为更好的诊断肝癌尤其是早期肝癌的血清标志物,因此GP73 的生理和病理作用受到广泛关注[13]。之前研究表明GP73 诊断临床肝癌的敏感性和特异性分别高达77%和93%,说明GP73可以作为早期肝癌诊断的标志物。早前我们实验室研究发现GP73 和CD206 的高表达与肝癌预后不良有关,现有研究也已经提供了关联GP73和致癌作用机制的一些见解,包括基质金属蛋白酶13(MMP13)的激活。此外,GP73 对肝癌的生长和转移具有免疫依赖性。GP73 可通过下调IL-12A 的表达使抗肿瘤淋巴细胞Th1 作用减弱。人Ad5E1A 基因作为腺病毒的极早期基因,具有抑癌基因的特性,携带E1A 基因而E1B 和E3 基因缺陷的腺病毒可靶向性在肿瘤细胞内增殖, 扩大杀伤肿瘤效果,同时又不会使原代细胞转化[14]。Ad5E1A 基因影响转录水平的表达,可以观察到当转入所构建的Ad5E1A 基因时,HepG2 细胞中GP73 的表达水平显著减低。进一步,我们发现与单独过表达GP73 相比,共同过表达Ad5E1A 和GP73 时肝癌细胞增殖水平被抑制。之前报道GP73 的表达水平与肝癌有密不可分的关系,然而两者之间的机制尚不明确。近年来靶向治疗成为热点,因此构建人Ad5E1A 基因表达载体,研究其与GP73 之间的关联,对于靶向治疗肝癌有重要的参考价值,可为临床治疗提供科学依据。
猜你喜欢
传染病信息(2022年3期)2022-07-15
中国动物保健(2022年2期)2022-05-05
太原理工大学学报(2021年6期)2021-11-25
海洋科学(2020年9期)2020-10-09
中国海洋大学学报(自然科学版)(2019年7期)2019-05-21
中国海洋大学学报(自然科学版)(2019年7期)2019-01-04
科技视界(2018年19期)2018-10-09
灾害医学与救援(电子版)(2018年1期)2018-06-05
上海农业学报(2017年4期)2017-04-10
安徽理工大学学报·自然科学版(2015年1期)2015-07-21
