
绿竹花发育相关BoAG-like基因的克隆与表达特性分析
2019-08-28 11:01魏涵天林新春
核农学报 2019年10期
魏涵天 时 燕 李 玲 林新春
(浙江农林大学省部共建亚热带森林培育国家重点实验室/浙江省竹资源与高效利用协同创新中心, 浙江 杭州 311300)
竹类植物属于禾本科竹亚科(Bambusoideae),是我国重要的经济植物。竹子开花会导致杆叶枯黄,地下茎逐渐变黑,失去萌发力,最终成片死去,严重影响生态环境和经济效益[1]。目前,已有关于竹类的研究报道,但竹子开花仍然是植物界的一个难解之谜[2-3],因此对竹子开花机理的研究具有十分重要的意义。
早在1991年,Coen等[4]提出了花发育ABC模型,认为存在着A、B和C三类同源异型基因,共同调控决定了花萼、花瓣、雄蕊和雌蕊四轮花器官的发育[5-8]。此后大量研究者对不同植株进行研究也发现,植物花器官的众多同源异型基因共同调控花器官的发育[9-11]。Angenent等[12]在此基础上,于1995年增加了参与调控胚珠的发育的D类基因,并将该模型发展为ABCD模型。Pelaz等[13]在2000年认为还需要E类基因参与调控四轮花器官,进一步将花发育模型发展为ABCDE模型。研究表明,竹子花发育为ABCDE模型,其中雷竹(Phyllostachysviolascens)的PpMADS1和PpMADS2[14]属于A类基因,绿竹(Bambusaoldhamii)的BoAP3[15]属于B类基因,绿竹的BoMADS1[16]属于C类基因,毛竹(Phyllostachysedulis)的PeMADS1[17]、雷竹的PvMADS5[18]和乌脚绿竹(Bambusaedulis)的BeMADS1[19]属于E类基因。
竹子C类基因(AGAMOUS,AG)属于MADS-box基因家族,调控雄蕊和雌蕊的发育[20]。目前,关于C类基因的报道并不常见,本研究以绿竹组培苗为试验材料,利用cDNA末端快速扩增技术(rapid-amplification of cDNA ends,RACE)获得了绿竹BoAG-like基因的cDNA全长;在生物信息学分析的基础上,对其表达模式进行研究,以期为进一步研究竹类花发育的分子调控机制奠定一定的理论基础。
1 材料与方法
1.1 材料
绿竹的花芽和营养植株根、茎、叶分别来源于浙江农林大学组培室继代一周后的花芽试管苗和营养试管苗[21]。采用Trizol法提取RNA,并于-70℃保存备用。
1.2 绿竹BoAG-like基因克隆与序列分析
采用3′RACE技术拼接出BoAG-like序列全长。根据3′-Full RACE Core set Ver.2.0试剂盒(TaKaRa,日本)引物设计原则,利用Primer 5.0设计了引物GSP1(5′-C A A C A G C G T G A A A G C A A C C A TT-3′)和GSP2(5′-T G A G C C A C A G G G A C T T A A G C A G CT-3′)。经1%琼脂糖检测后,纯化、回收PCR产物,并连接到pMD-20T载体;经热激法转化DH5α,在氨苯青霉素平板上进行蓝白斑筛选,挑取阳性克隆,并送至上海生工生物工程服务有限公司进行测序。测序结果经NCBI网站(https://www.ncbi.nlm.nih.gov/)上BLAST比对后,分别获取玉米(ZMM1、ZAG2、ZAG1、ZAGL5、ZMMADS4)、二穗短柄草(BdMADS18、BdAGL25、BdMADS3、BdMADS8、BdAGL6)、水稻(OsMADS58、OsMADS8、OsMADS6、OsMADS17)、毛竹(PeMADS3)、拟南芥(AtAGL1/SHP1、AtAGL5/SHP2、AtAG、AtAGL11/STK、AtAGL6)和杨树(PtAG、PtAGL11、PtAGL19、PtMADS1、PtAGL6)中的AG同源基因。利用DNAMAN软件对上述基因进行序列氨基酸编辑,并用MEGA 7软件构建系统进化树。
1.3 绿竹BoAG-like基因的组织特异性表达分析
以绿竹花芽试管苗不同时期的花芽和不开花试管苗的根茎叶为材料[22],分别提取RNA并反转录成cDNA;利用CFX96实时荧光定量分析仪(BIO-RAD,美国)对绿竹的花芽和根茎叶进行荧光定量分析。以Actin基因(登录号:BAC99043)为内参,Actin上游引物为5′-T G A G C T T C C T G A T G G G C A AG-3′,下游引物为3′-C C T G A T A T C C A C G T C G C A C TT-5′。BoAG-like上游引物为5′-G C C G T G G A C G T C T C T A C G AG-3′,下游引物为5′-T A G T G C T G G G C A G T G A T C T C TG-3′。荧光定量PCR反应程序:95℃预变性5 min;95℃变性15 s,60℃退火1 min,40个循环。采用2-△△Ct法[23]计算基因的相对表达量。
2 结果与分析
2.1 BoAG-like序列分析
通过前期的cDNA文库筛选工作发现,其中1条EST序列(Bo012)与AG基因序列相似性较高,可能与AG基因同源,且分析发现此序列缺少了3′端[21]。通过3′RACE技术扩增得到了该EST序列3′端的400 bp的片段,将3′RACE获得的序列与EST序列进行拼接,得到了1条全长为1 213 bp的cDNA序列,其含有1条完整的ORF序列,长度为792 bp。核苷酸序列比对结果发现,该序列为AG同源基因,与水稻、玉米、二穗短柄草等植物的AG基因有很高的同源性,故将其命名为BoAG-like。DNAMAN软件分析显示,该片段编码263个氨基酸,蛋白分子量约为100.36 kDa,等电点为5.00。Expasy protscale软件分析该基因蛋白的疏水性,结果显示其GRAVY值为-0.813,属于亲水性蛋白。
通过NCBI网站中的BLAST软件对比查找出各物种的AG基因及其同源基因,采用DNAMAN对这些基因编码的蛋白作序列比对。由图1可知,BoAG-like蛋白属于MADS-box家族,具有植物MADS-box蛋白典型的特征,其编码肽链包含MADS盒、K区2个保守区和Ⅰ区、C区2个非保守区,且具有AG基因特有的AG Ⅰ区和AG Ⅱ区;BoAG-like蛋白与来自其他单子叶植物的某些MADS-box蛋白高度同源,与水稻OsMADS58、玉米ZAG1、毛竹PhMADS58和二穗短柄草BdMADS18的一致性分别为89.41%、87.36%、89.31%和91.24%,与拟南芥AtAG和杨树PtAG的一致性分别为62.60%和64.73%。
在上述对比查找出来的蛋白中,挑选了几条和AG蛋白相似度比较高的序列,利用MEGA7软件中的邻接法(neighbor-joining,NJ)构建系统进化树。由图2可知,BoAG-like蛋白属于C类蛋白的eu-AG系,且该蛋白与水稻OsMADS58亲缘关系最近。
2.2 BoAG-like蛋白及其同源蛋白的二级、三级结构分析
蛋白质的二级结构主要有α-螺旋、β-折叠、β-转角等[23]。用SOMPA软件分析BoAG-like基因及其同源基因OsMADS58、BdMADS18、PhMADS3、ZAG1、AtAG、PLE和PtAGL11编码蛋白的二级结构(图3)。由表1可知,α-螺旋含量最高和最低分别为PhMADS3(60.75%)和AtAG(41.27%),无规则卷曲含量最高和最低分别为AtAG(32.14%)和PhMADS3(16.23%),BoAG-like的α-螺旋含量(55.51%)和无规则卷曲含量(20.91%)处于中等水平;延伸链含量最高和最低分别为OsMADS58(18.40%)和PLE(8.37%),β-转角含量最高和最低分别为PtAGL11(11.11%)和OsMADS58(6.25%),BoAG-like的延伸链含量(15.59%)和β-转角含量(7.98%)处于中等偏下水平。
蛋白质在形成立体三级结构时,其多肽链部分首先折叠形成α-型螺旋和β-型结构,再进一步折叠成立体球型从而发挥其生物学功能[24]。利用Swiss-model workspace对上述这8个蛋白的三级结构进行预测,结果发现BoAG-like蛋白与OsMADS58蛋白、BdMADS18蛋白相似度最高(图4)。这三者蛋白都具有相似的同源二聚体(homo-dimer)和同源四聚体(homo-tetramer),其中OsMADS58蛋白和BdMADS18蛋白空间构型更为接近,然而BoAG-like蛋白与OsMADS58和BdMADS18两个蛋白在同源二聚体和同源四聚体的空间结合上又存在明显的差异。
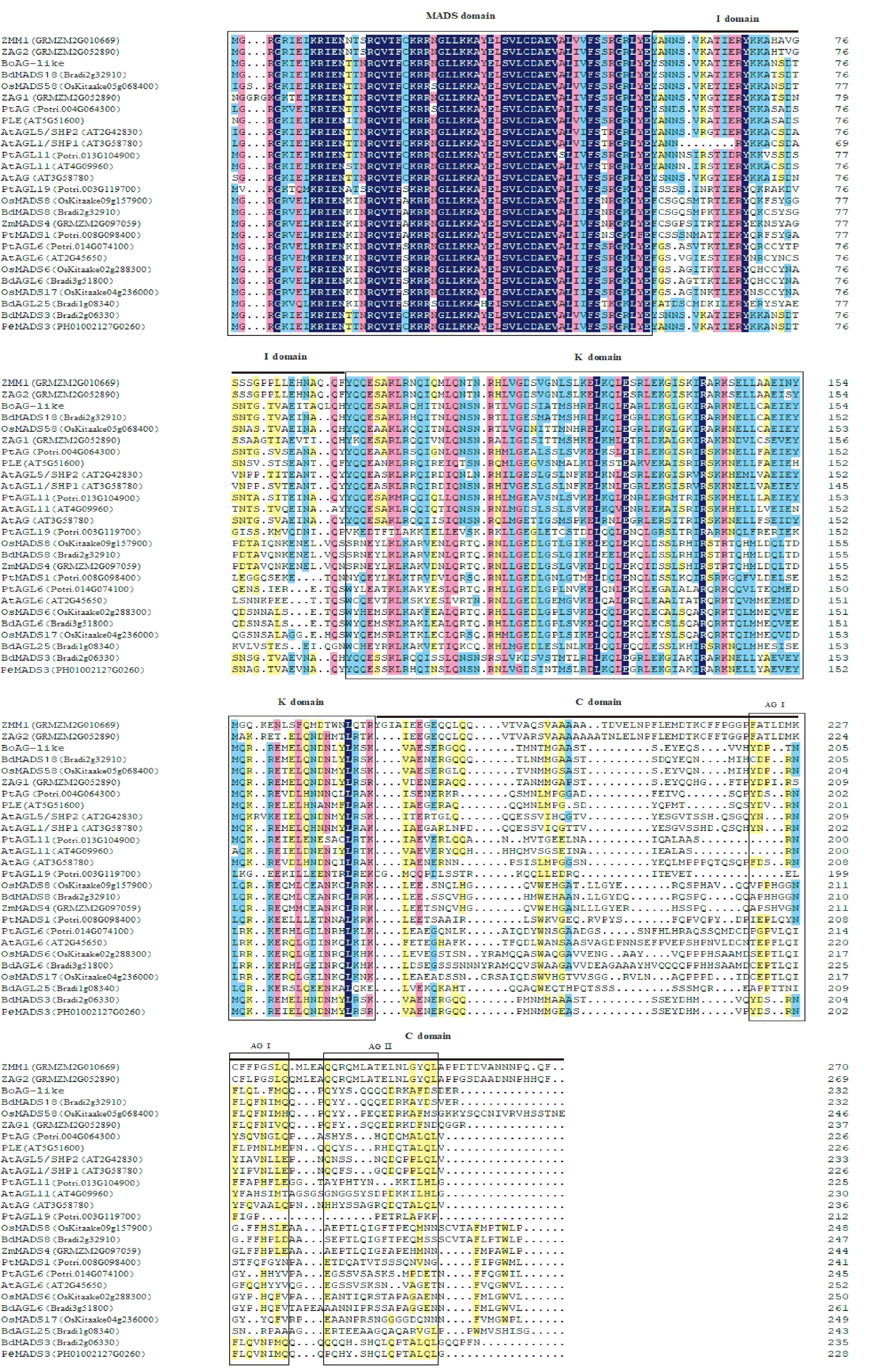
图1 BoAG-like与其他物种相关MADS蛋白序列比较Fig.1 Amino acid sequence comparison of BoAG-like and the related MADS proteins
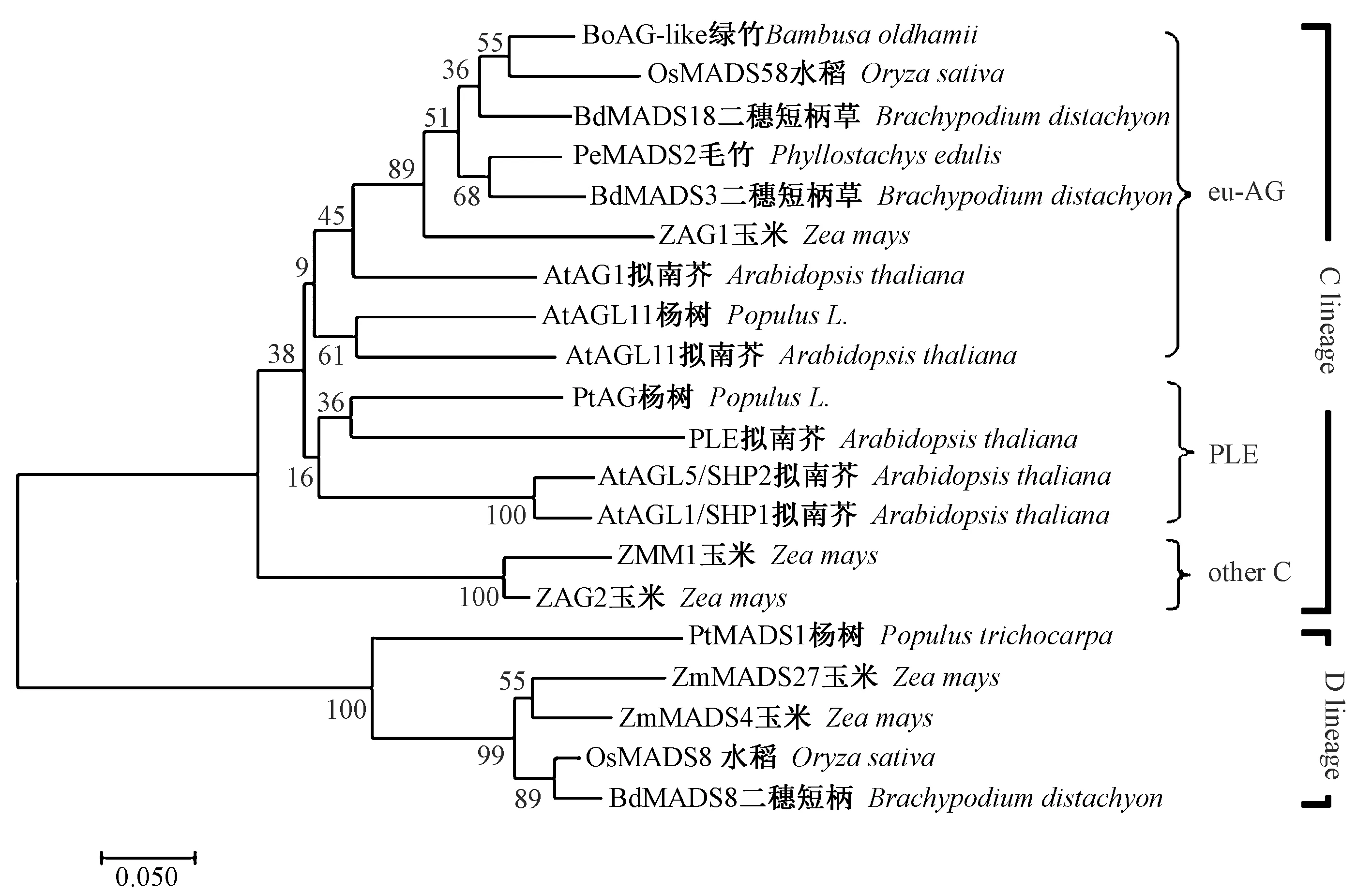
图2 基于BoAG-like基因氨基酸序列构建的同源基因系统进化树Fig.2 Phylogenetic analysis of homologous genes based on the amino acid sequence of BoAG - like

图3 BoAG-like蛋白及其同源蛋白的二级结构的分布Fig.3 Predicted secondary structure of BoAG-like protein and its’ homologous proteins
2.3 BoAG-like组织特异性分析
由图5可知,荧光定量PCR结果表明BoAG-like在花器官中表达量最高,在营养器官中的表达量都较低。其中BoAG-like在根、茎、叶中的表达量均较低,在花芽中的表达量最高(图5-A);绿竹花发育过程中,BoAG-like在花芽中的表达量呈逐渐升高的趋势,在花芽生长后期达到顶峰(图5-B);且不同花芽部分中,BoAG-like在雌蕊中的表达量最高,其次为雄蕊,浆片再次之,内稃中表达量较低,外稃中的表达量最低,且雌蕊中的表达量明显高于其他花芽部位中的表达量(图5-C)。表明BoAG-like在绿竹花发育过程起到了重要的作用。对不同发育时期的绿竹花芽进行解剖,观察发现绿竹花器官由内稃和外稃包被,内含浆片,由3~4个雄蕊和1个雌蕊组成(图6)。图7为不同时期的绿竹大中小花芽。
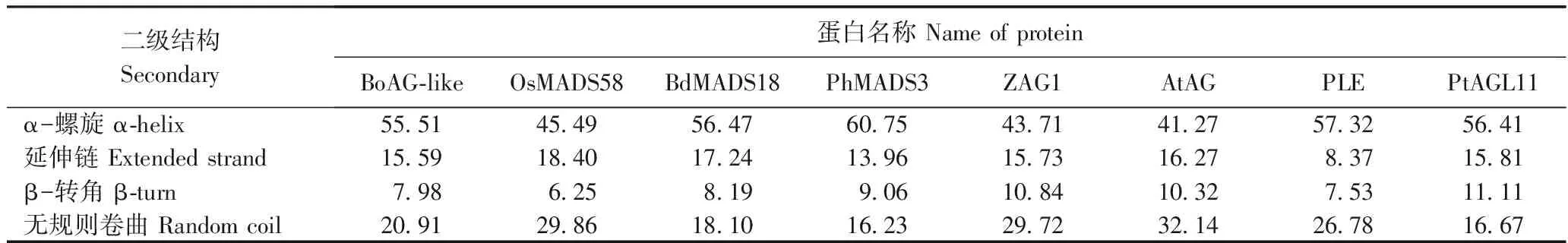
表1 BoAG-like蛋白及其同源蛋白的二级结构元件组成Table 1 Composition of predicted secondary structure of BoAG-like and its homologous proteins/%
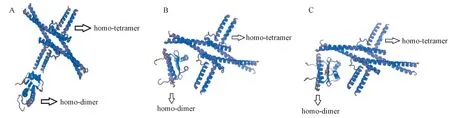
注:A:绿竹BoAG-like蛋白三级结构预测图;B:水稻OsMADS58蛋白三级结构预测图;C:二穗短柄草BdMADS18蛋白三级结构预测图。Note: A: Predicted tertiary structures of BoAG-like of Bambusa oldhamii. B: Predicted tertiary structures of OsMADS58 of Oryza sativa. C: Predicted tertiary structures of BdMADS58 of Brachypodium distachyon.图4 BoAG-like蛋白及其相似蛋白的三级结构预测图Fig.4 Predicted tertiary structures of BoAG-like and its’ similar proteins
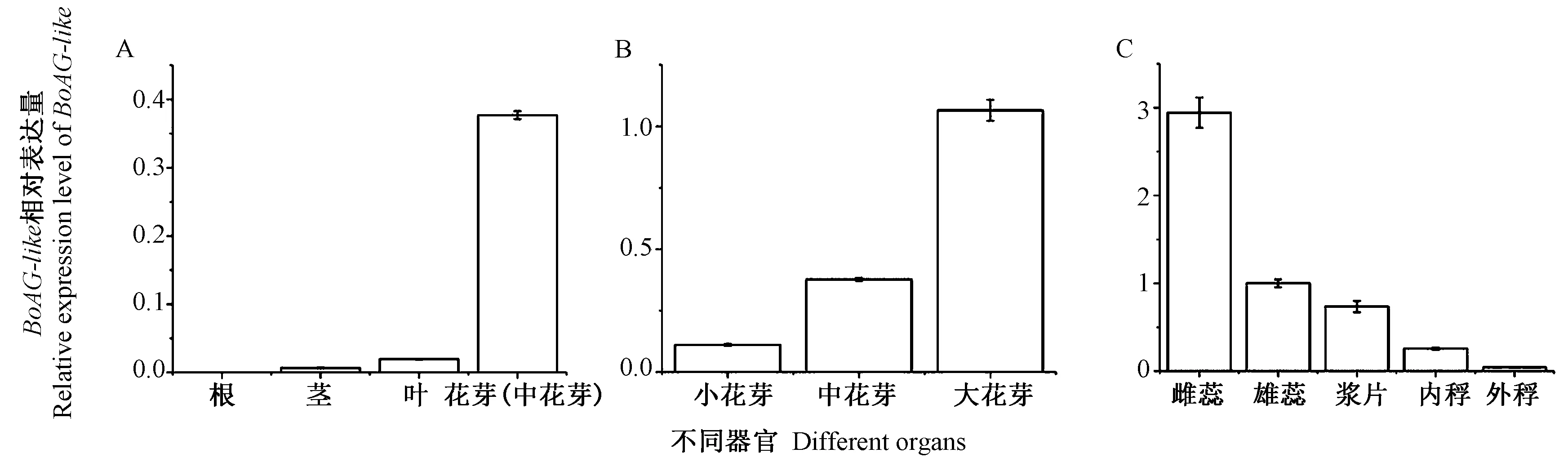
注:A: 不同组织; B:花发育过程; C: 不同花芽部分。Note: A: Different tissues. B: Flower development stages. C: Different flower buds partial.图5 BoAG-like基因的表达模式Fig.5 The expression pattern of BoAG-like in Bambusa oldhamii

注:Pa:内稃,Le:外稃,Lo:浆片,An:雄蕊,Pi:雌蕊。Note: Pa: Palea. Le: Lemma. Lo: Lodicule. An: Androecium. Pi: Pistil.图6 绿竹花芽解剖图Fig.6 The anatomical morphology of flower bud in Bambusa oldhamii

图7 不同时期的绿竹花芽Fig.7 Flower buds of Bambusa oldhamii at different developmental stages
3 讨论
AG基因家族进化过程中产生了euAG和PLE两个进化系[25]。在拟南芥中C功能基因AtAG属于euAG类,而在金鱼草(Antirrhinummajus)中,C类功能基因AmPLE属于PLE类[26]。本研究分离的BoAG-like蛋白质序列在系统进化树中属于C类基因的euAG系,BoAG-like与水稻、二穗短柄草等AG-like同源蛋白所编码的氨基酸序列同源性达到80%以上,且在系统进化树上聚为一类。在花发育的ABC模型中,C类基因在第3轮器官和B类共同作用形成雄蕊,在第4轮器官单独作用形成雌蕊[27-28]。荧光定量PCR结果表明,BoAG-like随着花发育的过程表达量呈上升趋势,在花芽中BoAG-like的雌蕊和雄蕊表达量最高,推断该基因可能参与了花器官的发育。
绿竹BoAG-like和水稻OsMADS58及二穗短柄草BdMADS18序列一致性较高,分别为89.41%和91.24%,这三者编码蛋白的二级结构元件组成也十分相似,预测得到的蛋白质三级结构均具有相似的同源二聚体和同源四聚体,但OsMADS58蛋白和BdMADS18蛋白的三级结构更为接近,BoAG-like蛋白的三级结构与这两个蛋白在同源二聚体和同源四聚体的空间结合上存在着明显的差异。此外,绿竹BoAG-like、水稻OsMADS58和二穗短柄草BdMADS18均属于C类基因,控制着雄蕊和雌蕊的发育,但绿竹BoAG-like基因在花芽中雌蕊的表达量远高于雄蕊;而水稻OsMADS58在花发育初期仅在雄蕊中有表达,在花发育的中后期OsMADS58逐渐在心皮中有表达,但在整个花发育过程中OsMADS58在雌蕊的相对表达量均最高[29];二穗短柄草BdMADS18在雄蕊、心皮、浆片和内稃中均有明显的表达,在雌蕊中的表达量最高[30]。绿竹BoAG-like与水稻和二穗短柄草的同源蛋白的蛋白质结构存在差异,其原因可能与该基因特殊的表达模式有关,也可能与竹子独特的开花生物学特性有关。
4 结论
本试验克隆获得了绿竹BoAG-like,该基因具有AG基因特有的AG Ⅰ和AG Ⅱ区。系统进化树表明BoAG-like属于C类基因的eu-AG系,且该基因编码的蛋白与水稻OsMADS58和二穗短柄草BdMADS18亲缘关系最近。蛋白三级结构预测表明,BoAG-like与OsMADS58和BdMADS18最为相似,但也存在明显差异。荧光定量PCR结果表明,BoAG-like基因在花器官的表达量远高于营养器官的表达量,且随着绿竹花发育的进程呈上升趋势;BoAG-like表达量在绿竹花芽不同部位存在明显差异,其中在雌蕊中最高,在内稃和外稃中的表达量较低,表明该基因可能参与了竹子花器官的发育。本研究结果为阐明竹子花发育的分子机制奠定了理论基础。下一步可通过纯化蛋白,对BoAG-like蛋白进行空间结构解析,并采用过表达和基因编辑技术对BoAG-like基因进行功能验证。
猜你喜欢
经济林研究(2022年2期)2022-07-20
课外生活·趣知识(2022年6期)2022-07-14
现代农业科技(2022年4期)2022-03-08
学生天地(2020年15期)2020-08-25
山西农业科学(2018年9期)2018-09-12
文苑·经典美文(2018年6期)2018-06-23
生物学教学(2017年6期)2017-02-18
红蜻蜓·低年级(2016年9期)2016-05-14
传奇故事(破茧成蝶)(2015年7期)2015-02-28
火花(2015年8期)2015-02-27
