
大花耧斗菜雄蕊发育过程研究
2018-09-12 02:25王金耀宋红贤邢国明亢秀萍
山西农业科学 2018年9期
王金耀 ,杨 阳 ,,宋红贤 ,李 森 ,邢国明 ,亢秀萍
(1.山西农业大学园艺学院,山西 太谷 030801;2.吕梁学院生命科学系,山西 吕梁 033000)
大花耧斗菜(Aquilegia glandulosa)是毛茛科耧斗菜属的宿根花卉,其花型独特如漏斗,花大而繁多,花色亮丽,花期长且耐寒,有很高的观赏价值[1-2]。耧斗菜展现出广泛的生态学和形态学多样性,保持了较高的种间杂交亲和性[3-4],因经,开展耧斗菜的生理和形态的功能遗传学分析,对被子植物进化研究具有很大的价值[5]。此外,退化雄蕊不能产生可育花粉,不能发挥雄性生殖的作用,但是在进化过程中,一部分退化雄蕊获得了一些新功能[6],帮助植物繁殖,研究不同的近缘关系的物种间退化雄蕊的功能差异,可揭示退化雄蕊在被子植物系统进化中的意义[7-8]。
本研究利用常规石蜡切片技术,从显微水平上观察了大花耧斗菜低温春化后雄蕊原基分化及发育的过程,掌握大花耧斗菜的生殖发育规律,以期为耧斗菜退化雄蕊作用的研究提供借鉴。
1 材料和方法
1.1 试验材料
供试材料为大花耧斗菜(Aquilegia glandulosa),种子购买于泛美种子公司。
1.2 试验方法
2015年1月在山西农业大学日光温室苗床播种大花耧斗菜种子,待幼苗长至12片叶时,放置于人工气候箱为5℃低温春化处理,处理结束将植株放置在日均温为20℃的自然条件下生长,每3 d取材一次。取材后剥去外层莲座叶,露出小叶包裹的茎尖,立即放入FAA固定液中固定并保存[9],常规石蜡切片,采用番红-固绿双重染色法[10-11]在低倍显微镜下进行观察。
2 结果与分析
2.1 大花耧斗菜雄蕊的形态结构
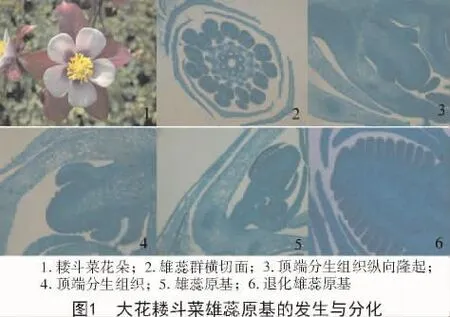
耧斗菜花呈辐射对称,由外到内依次是5枚萼片、5枚花瓣和雄蕊群(图1-1)。雄蕊多数,共10轮,每轮有5枚雄蕊,其中5轮雄蕊与萼片对生,5轮与花瓣对生,显示出雄蕊群轮间交互(图1-2)。雄蕊群外部的花丝短,花药较大;靠近内侧花丝伸长,花药逐渐变小。退化雄蕊的花丝最长,持续时间最长,花药最小。花丝表皮光滑,圆柱形。花药底生,呈黄色椭圆形,表面有明显的分界,分为左右两半。
2.2 雄蕊原基的发生与分化
大花耧斗菜的花原基分化出花瓣原基后,顶端分生组织纵向高度隆起呈球状(图1-3),在球状分生组织底部、花瓣原基内侧,对称出现横向隆起,分化成最外轮雄蕊原基。隆起的组织呈圆柱状,细胞小,细胞大小相同,细胞质浓密(图1-4)。
最外轮雄蕊原基发生后,雄蕊原基继续横向分化生长。其内侧的雄蕊原基依次向顶端发生,可以看出雄蕊原基的发育顺序是由外侧向内侧,即最外部雄蕊原基首先分化,然后依次产生其他每一轮雄蕊原基,直到最后一个退化雄蕊原基出现。先发育的外侧雄蕊原基最大,依次发育的内侧雄蕊原基逐轮变小,退化雄蕊原基最小(图1-5、图1-6)。
2.3 花丝的发育
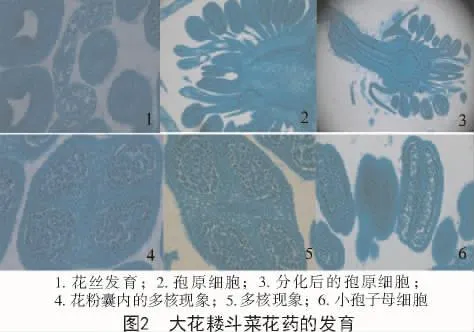
大花耧斗菜雄蕊原基分化结束后,进入了雄蕊的发育阶段,每一个圆柱形的雄蕊原基顶端膨大发育成花药,下端发育成花丝,花丝的结构简单,最外层是一层角质化的表皮细胞,表皮内是薄壁细胞,中央由维管束贯穿(图2-1),直达药隔,为花药的发育提供营养物质。从雄蕊群的花丝发育来看,最顶端的退化雄蕊的花丝生长发育早于下端雄蕊的花丝发育,并且从花丝长度来看,上端雄蕊的花丝长度均长于下端的雄蕊花丝。退化雄蕊的花丝在发育最后阶段,中央维管束消失,只剩下角质化的表皮细胞,其他功能雄蕊的花丝不存在上述现象。
2.4 花药的发育
全部雄蕊原基分化结束后,顶端即最内层雄蕊顶端最先开始膨大,最下端及最外层的雄蕊发育明显滞后。从切片上可以看出,雄蕊的发育顺序是从顶端至下端,最内侧向外侧离心式发育。圆柱形雄蕊顶端膨大发育,最后发育成椭圆形花药。从花药的横切和纵切可以看出,未成熟大花耧斗菜的花药分为4个药室,纵轴长于横轴,且花药中央有很明显的分界。
雄蕊发育早期,雄蕊顶端外有一层表皮,表皮层内是一群形状相似、结构简单、分裂活跃的幼嫩细胞即花药原始体。从花药的纵切面看,花药膨大,花药中央出现细胞质较浓,染色较深,呈现一列或数列排布的孢原细胞(图2-2)。孢原细胞经过分裂和分化,外层细胞与表皮一起构成花粉囊的壁层,内层细胞为造孢细胞。花药中部细胞进一步分裂、分化,出现了药隔和长椭圆形的维管束。
花粉囊最内层为绒毡层,大花耧斗菜绒毡层的细胞比外围的壁细胞大,多为长方形或多边形,紧密的排列在花粉囊的内壁,具有多核现象(图2-4,5)。随着花药的发育,绒毡层细胞开始分解消失。切片观察中发现,退化雄蕊也会形成较小的花粉囊,但花粉囊内层绒毡层细胞与正常功能雄蕊不同。
造孢细胞经过分裂,形成大量小孢子母细胞,大花耧斗菜小孢子母细胞为圆球形,细胞体积大,原生质浓厚。小孢子母细胞进一步发育,形成圆球形小孢子(图2-6)。
2.5 退化雄蕊
在对耧斗菜花部结构的分化发育观察中发现,耧斗菜的退化雄蕊原基位于雄蕊群原基顶端,退化雄蕊是在最后一轮功能雄蕊原基发生后发生,之后生长锥顶端变平,开始分化雌蕊原基。说明耧斗菜雄蕊群原基的发生是向心式发育。但雄蕊原基发生后,进入小孢子发育时期,观察到退化雄蕊发育先于下部的功能雄蕊,最后会形成在显微镜下观察到的成熟花粉粒。在退化雄蕊发育过程中,起初花丝的发育是正常的,具有正常的结构,但是在发育中期,维管束和薄壁细胞停止生长,并且慢慢消失,花丝结构只剩下最外层的表皮细胞,花药中未形成和功能雄蕊一样的绒毡层结构,同时花粉母细胞体积小,花粉囊较小,并在切片中看到形成了小孢子(图2-3)。
3 结论与讨论
大花耧斗菜花呈辐射对称,最外层是5片萼片,其次是5片花瓣,内侧是雄蕊群,雄蕊多数,共10轮,每轮有5枚雄蕊,其中5轮雄蕊与萼片对生,其余5轮与花瓣对生,从而显示出雄蕊群轮间的交互[12]。雄蕊原基的发育顺序是由外侧向内侧,即最外部雄蕊原基首先发育,然后依次产生其他每一轮雄蕊原基,直到最后一个退化雄蕊原基出现[13]。因此,耧斗菜花的雄蕊原基的发生属于向心式发生[14]。从雄蕊群的花丝发育来看,最顶端的退化雄蕊的花丝生长发育早于下端雄蕊的花丝发育,并且从花丝长度来看,上端雄蕊的花丝长度均长于下端的雄蕊花丝[15]。退化雄蕊的花丝在发育最后阶段,中央维管束消失,只剩下角质化的表皮细胞,其他功能雄蕊的花丝不存在上述现象。
雄蕊原基分化结束后,顶端及最内层雄蕊顶端最先开始膨大,最下端及最外层的雄蕊发育明显滞后。从切片上可以看出,雄蕊的发育顺序是从顶端至下端,及最内侧向外侧,离心式发育[16]。圆柱形雄蕊顶端膨大发育,最后发育成椭圆形花药[17-18]。从花药的横切和纵切中可以看出,未成熟大花耧斗菜的花药分为4个药室,纵轴长于横轴,且花药中央有很明显的分界。
在对耧斗菜花部结构的分化发育观察中发现,耧斗菜的退化雄蕊原基位于雄蕊群原基顶端,退化雄蕊是在最后一轮功能雄蕊原基发生后发生,之后生长锥顶端变平,开始分化雌蕊原基,可以看出,耧斗菜雄蕊群原基的发生是向心式发育。但雄蕊原基发生后,进入小孢子发育时期后,观察到退化雄蕊发育先于下部的功能雄蕊,最后会形成在显微镜下观察到的成熟花粉粒。在退化雄蕊发育的过程中,起初花丝的发育是正常的,具有正常的结构,但是在发育中期,维管束和薄壁细胞停止生长,并且慢慢消失,花丝结构只剩下最外层的表皮细胞,花药中未形成和功能雄蕊一样的绒毡层结构,同时花粉母细胞体积小,花粉囊较小,并在切片中看到形成了小孢子。退化雄蕊是否可以形成正常的花粉粒,且是否有活力,还需进一步探索。
猜你喜欢
现代园艺(2023年3期)2023-02-01
工会博览(2022年17期)2022-07-15
华北农学报(2020年1期)2020-04-16
花生学报(2019年2期)2019-10-22
计算机应用(2018年10期)2018-11-22
华南师范大学学报(自然科学版)(2017年4期)2017-09-11
中华手工(2017年6期)2017-07-06
浙江大学学报(农业与生命科学版)(2017年2期)2017-05-19
生物学教学(2017年6期)2017-02-18
中华手工(2016年7期)2016-10-10
