
接种贝莱斯芽孢杆菌SW5菌株对发酵鳀鱼鱼露品质的影响
2019-08-28 11:02杨海宁宁豫昌王昌毓翁佩芳吴祖芳朱亚珠
核农学报 2019年10期
杨海宁 宁豫昌,2 王昌毓 翁佩芳 吴祖芳,* 朱亚珠
(1 宁波大学食品与药学学院,浙江 宁波 315832;2 河南牧业经济学院,河南 郑州 450046;3 浙江国际海运职业技术学院, 浙江 舟山 316021)
鱼露又称鱼酱油,是一种以海洋鱼类蛋白质水解制得的汁液,色泽呈琥珀色,味道鲜美且带有咸味和鲜味。鱼露富含多种营养物质,除含有人体所需的8种必需氨基酸外,还含有多种有机酸和微量元素[1],在一些地区鱼露甚至还作为一种必需膳食蛋白和氨基酸的来源[2]。长期以来,鱼露的生产以自然发酵法为主,但该方法存在发酵周期过长(通常1~2年)等缺陷,制约了鱼露产业的发展。目前,利用外加微生物促进鱼蛋白降解已成为缩短鱼露发酵周期的重要研究方向。Yongsawatdigul等[3]向印度鳀鱼中接种分离于自然发酵鱼露中的蛋白酶产生菌枝芽孢杆菌(Virgibacillussp)SK37,发现经发酵4个月的鱼露总氨基酸含量与传统发酵12个月的样品相当。Akolkar等[4]报道采用蛋白酶产生菌-嗜盐菌(Halobacteriumsp.)SP1(1)作为起始培养物可以加速鱼露发酵,且所获鱼露在必需氨基酸和风味氨基酸含量方面优于商品鱼露。Udomsil等[5]报道向印度鳀鱼中按顺序添加分离自发酵鱼露中的嗜盐四联球菌(Tetragenococcushalophilus)MS33和蛋白酶产生菌枝芽孢杆菌(Virgibacillussp.)SK37,不仅可以提高鱼露中挥发性化合物和谷氨酸的含量,还能提高鱼露的整体可接受性。
甲基营养型芽孢杆菌(Bacillusmethylotrophicus)是2010年发现的新菌种[6]。通过比较基因组学结合形态、生理、化学分类和系统发育学分析,已证实甲基营养型芽孢杆菌KACC 13015T与贝莱斯芽孢杆菌(Bacillusvelezensis) NRRL B-41580T具有表型一致性和基因型一致性[7],即甲基营养型芽孢杆菌同归属于贝莱斯芽孢杆菌。曹红等[8]在研究贝莱斯芽孢杆菌时,发现该菌种能产生高活力胞外蛋白酶;此外,研究表明,该菌种在适宜条件下可产生淀粉酶[9]、纤维素酶[10]、几丁质酶[11]和脂肪酶[12]等。由于该菌种具有良好的生物安全性[13]和益生功能[14],且该菌株具有一定的耐盐性,因此其具有应用于食品发酵的潜力。但目前尚未见到将该菌种应用于鱼露发酵的相关报道,故本研究以低值鳀鱼为原料,接种SW5菌株,在发酵期间测定鱼露营养价值的重要指标氨基酸态氮和水产品新鲜度主要指标挥发性盐基氮等,在发酵结束后对发酵醪液挥发性风味物质进行检测,以判断接种贝莱斯芽孢杆菌SW5菌株对发酵鳀鱼鱼露品质的影响。
1 材料与方法
1.1 材料
贝莱斯芽孢杆菌SW5,由宁波大学食品生物技术实验室保藏;冷冻鳀鱼,购自浙江舟山兴业有限公司;舟山商品鱼露,购自浙江舟山兴业有限公司。
1.2 主要仪器与设备
7890B7000C气相色谱质谱联用,美国安捷伦公司;SPME 75 μm萃取头,美国Supelco公司;PHS-3C型pH计,上海圣科仪器设备有限公司;TE214S分析天平,成都浩驰仪器有限公司;XPX智能生化培养箱,江南仪器厂;SW-CJ-2D型超净工作台,苏州净化设备有限公司;LDZX-40B Ⅰ型立式蒸汽灭菌器,上海申安医疗器械厂;5804R高速大容量冷冻离心机,德国Eppendorf公司;MGH-90电动搅肉机,佛山名健电器制造有限公司;QYC-2102C全温振荡培养箱,宁波江南仪器厂。
1.3 试验方法
1.3.1 种子液制备 将SW5菌株从试管斜面接种至40 mL/100 mL的营养肉汤中,37℃、120 r·min-1恒温振荡培养12 h,备用。
1.3.2 SW5菌株生长曲线和产酶曲线测定 将培养12 h的贝莱斯芽孢杆菌SW5以5%的接种量加入100 mL/250 mL的营养肉汤中,37℃、120 r·min-1振荡培养,每隔2 h取一次样品,采用比浊法[15]测定SW5菌株的生长曲线。以国标GB/T 23527-2009[16]中福林酚法测定SW5菌株蛋白酶活力,每个样品测定3次,并绘制SW5菌株产酶曲线。
1.3.3 原料前处理 将冷冻鳀鱼流水解冻,沥干水分后去除头和内脏。将鳀鱼与蒸馏水按照固液比1∶0.6(m/v)混合并使用绞肉机打成鱼糜[17],然后装入500 mL锥形瓶中,每瓶200 g,放入75℃水浴锅中,待鱼糜样品中心温度达到75℃后保温21 min[18],以达到巴氏消毒的目的,冷却至室温,备用。
1.3.4 鱼露样品制备 试验分组:处理组1:取5%鱼糜总量的贝莱斯芽孢杆菌种子液,8 000 r·min-1离心(4℃)15 min得到菌体细胞,然后将菌体细胞接种至巴氏消毒后的鱼糜中,37℃、120 r·min-1摇床培养3 d,添加20%食盐,继续发酵4 d;处理组2:巴氏消毒后的鱼糜除了不接种试验菌,其余操作均与处理组1相同;处理组3:将解冻鳀鱼(不去除头和内脏)与蒸馏水按照固液比1∶0.6混合并使用绞肉机打成鱼糜,直接加入20%食盐混匀,模拟自然发酵。每个处理组均作3次重复。
分析样品制备:取20 mL发酵醪液,4℃、8 000 r·min-1离心10 min,取中层清液,重复离心(4℃,8 000 r·min-1,10 min)1次,合并2次离心得到的中层清液,双层滤纸过滤后制备成待测样品。
1.3.5 理化指标测定 氨基酸态氮(amino acid nitrogen, AA-N)含量测定采用甲醛滴定法[19];挥发性盐基氮(total volatile basic nitrogen, TVB-N)含量测定采用半微量定氮法[19];pH值测定采用数字式pH计;总酸含量测定采用甲醛滴定法。
1.3.6 GC-MS分析挥发性风味成分 吸取3 mL待测样品放入标准顶空进样小瓶内,其气相部分(顶空)导入顶空进样装置连接的气相色谱。萃取头老化:按照SUPELCO公司推荐的条件,第一次使用时将50/30 μm DVB/CAR/PDMS萃取头在气相色谱进样口210℃老化1 h,之后每次使用前老化0.5 h,至无干扰峰出现。
顶空固相微萃取:准确取3 mL待测样品,置于顶空瓶中,于60℃平衡15 min。将活化好的固相微萃取器插入平衡后的样品,顶空恒温振荡萃取30 min,振荡速率为250 r·min-1。取出插入气相色谱柱的进样口解吸附7 min,解析温度为210℃。
色谱条件:毛细管柱为Vocal(60 m×0.32 mm×1.8 μm)。载气为高纯氦气,恒定流速3 mL·min-1,进样口温度210℃,不分流进样模式。柱温箱采用程序升温:起始温度35℃,保持3 min,以3℃·min-1升至40℃,保持1 min,再以5℃·min-1升至200℃,保持20 min。
质谱条件:采用全扫描模式采集信号,电离方式EI,电子轰击能量为70 eV;接口温度220℃,离子源温度230℃,四级杆温度150℃,扫描质量范围m/z 40~600,扫描频率3.6 scans·s-1。
定性定量分析:化合物经计算机检索与NIST 14 Library相匹配,选择较高匹配度的检索结果与质谱图库中的标准谱图对照、复合,进行人工谱图解析确认挥发性成分,并结合文献报道的已知化合物确认检测物成分。按照面积归一化法进行定量分析,求得各化学成分的相对百分含量[20]。将舟山商品鱼露与上述待测样品一同测量,并进行比较分析。
1.3.7 感官评价分析 采用描述性定量分析(quantitative descriptive analysis test)法对样品进行感官分析,评定小组由10位食品专业的老师与研究生组成(5男5女,年龄在20~35岁之间)。感官评定前,对评定人员进行一致的鱼露感官特性描述培训,培训后对鱼露的感官特性鲜味(umami)、腐臭味(rancid)、鱼腥味(fishy)、酸味(sour)、肉味(meaty)、咸味(salty)、干酪味(cheesy)、水果香味(fruity)进行打分,计分采用5分制,0分表示该风味不能被感受到,5分代表该风味非常强烈。
1.4 数据分析
每个样品平行测定3次,所有数据采用SAS 8.1软件分析,应用单因素方差分析(One-way ANOVA)和邓肯法(Duncan’s)对试验数据进行差异分析。采用OriginPro 9.1制图。
2 结果与分析
2.1 SW5菌株生长曲线和产酶曲线
由图1可知,SW5菌株在该培养条件下无延滞期,对数期是0~16 h,稳定期是16~26 h,衰亡期是26~48 h,在后续试验中种子菌龄统一选择培养时间为12 h的。由SW5菌株产酶曲线可知,该菌株从2 h开始产生蛋白酶,26 h和32 h有2个产酶高峰,至48 h发酵液中仍有高活力蛋白酶,说明该菌株可以用于发酵制备鱼露。
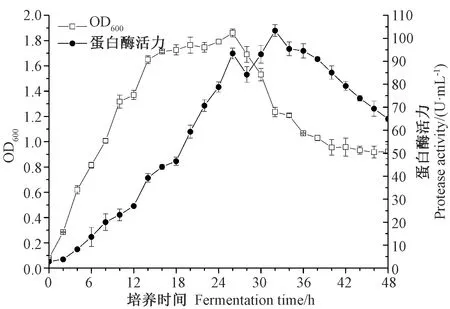
图1 SW5菌株生长曲线和蛋白酶活力曲线Fig.1 Growth curve and protease activity curve of strain SW5
2.2 发酵过程中理化指标结果分析
2.2.1 发酵过程中氨基酸态氮含量 氨基酸态氮(AA-N)是以氨基酸形式存在的氮元素含量,该指标越高,说明鱼露中氨基酸含量越高,是衡量鱼露品质的重要指标。由图2可知,随着发酵时间的延长,处理组2 AA-N含量几乎不变,处理组3 AA-N含量缓慢增加。前期预试验表明处理组1 AA-N含量至第6天时达到最大值,为0.76 g·100 mL-1,达到市售二级鱼露水平[21],随后出现下降趋势。整个发酵过程中处理组1中AA-N始终高于处理组2和处理组3,可能是SW5菌种产生的大量蛋白酶可将处理组1中鱼蛋白酶解,从而导致AA-N含量快速上升。综上,在接种该菌株6 d时,发酵醪液中AA-N含量最高。后续试验以此为依据,选择6 d作为发酵周期。
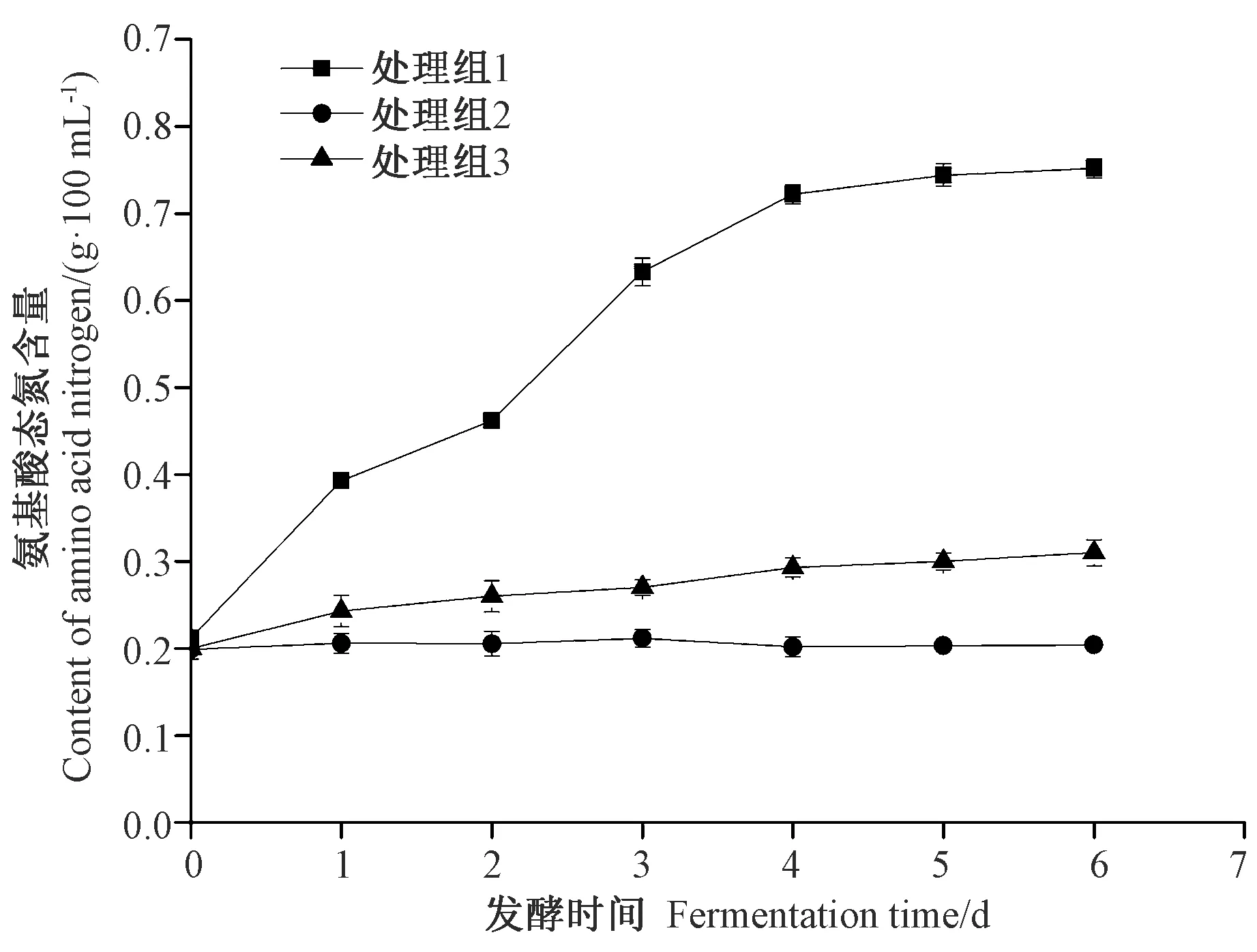
图2 发酵过程中氨基酸态氮含量的变化Fig.2 Changes of amino acid nitrogen content during fermentation

图3 发酵过程中挥发性盐基氮含量的变化Fig.3 Changes of total volatile basic nitrogen content during fermentation
2.2.2 发酵过程中挥发性盐基氮含量 挥发性盐基氮(TVB-N)指蛋白质、氨基酸等在微生物作用下分解形成氨、三甲胺等碱性含氮化合物,是衡量鱼露腐败变质的重要指标。由图3可知,随着发酵时间的延长,处理组2 TVB-N含量略有增加,但变化不明显;处理组3 TVB-N含量呈现逐渐增加趋势,至发酵第6天时,发酵醪液TVB-N含量为57.63 mg·100 mL-1,高于处理组2;而处理组1 TVB-N含量在发酵第1天缓慢增加,1~4 d快速增加,4 d后增加趋势变缓,至发酵第6 天时,发酵醪液TVB-N含量为104.53 mg·100 mL-1,高于处理组2和处理组3。处理组1TVB-N含量持续增加可能与接种SW5菌株有关,SW5菌株在产生蛋白酶外可能还产生其他酶,将发酵醪液中氨基酸继续分解为一些含氮化合物,导致TVB-N含量升高。
2.2.3 发酵过程中pH值和总酸的变化 鱼露pH值和总酸是由游离氨基酸、有机酸类物质和含氮化合物分解产生的微碱性物质共同作用的结果。由图4可知,随着发酵时间的延长,处理组1 pH值先升高后下降,至发酵第6天时,发酵醪液pH值为5.86;处理组2与处理组3 pH值呈缓慢下降趋势,至发酵第6天时,发酵醪液pH值分别达到5.39和5.41。由图5可知,整个发酵过程中,处理组1和处理组2总酸含量几乎不变,可能是由于SW5菌株产酸能力较弱;而处理组3发酵醪液总酸含量呈上升趋势,至发酵第6天时,其总酸含量达到0.91 g·100 mL-1。
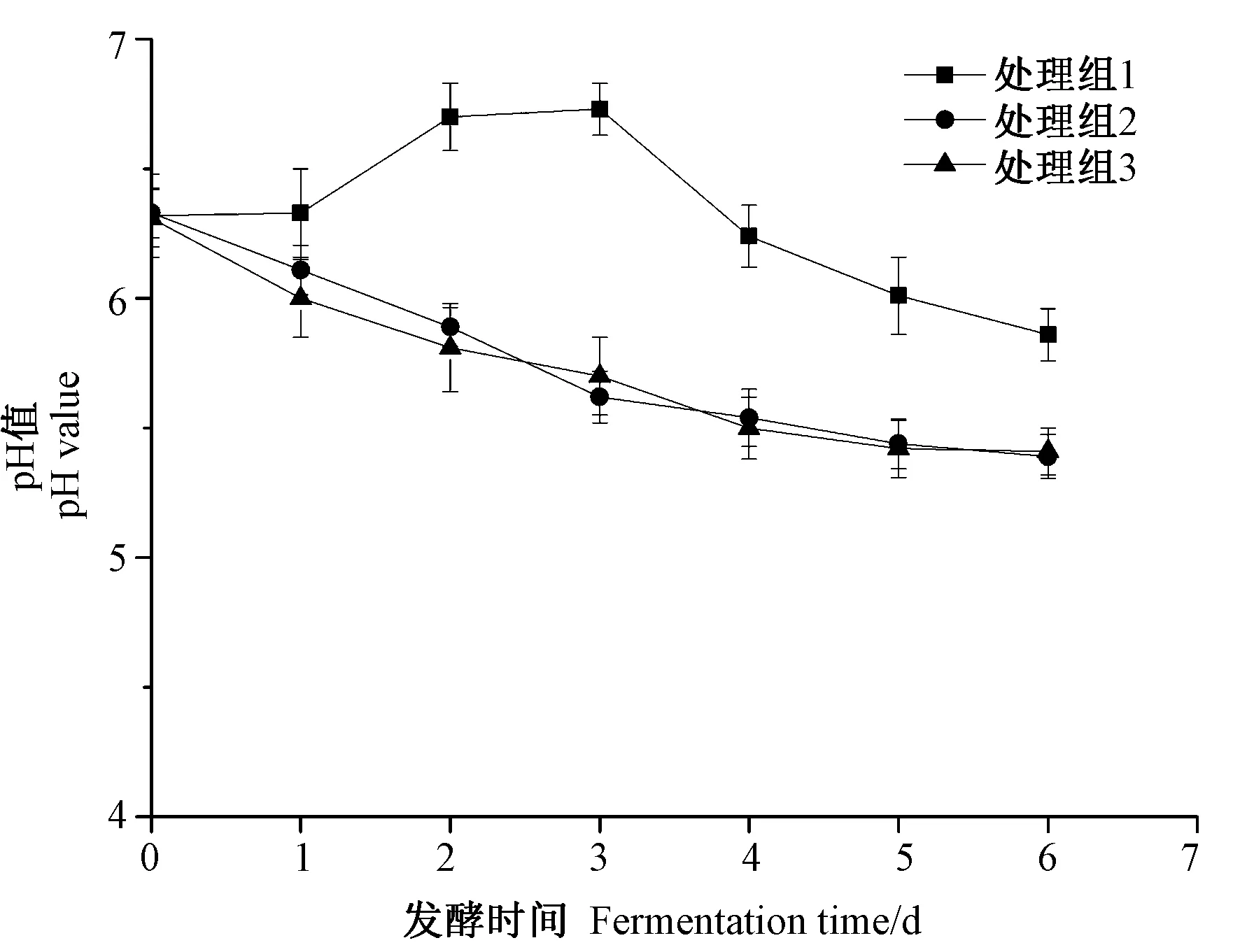
图4 发酵过程中pH值的变化Fig.4 Changes of pH value during fermentation
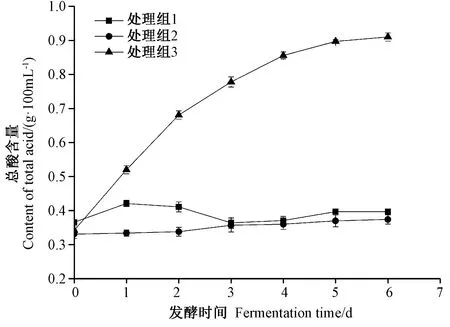
图5 发酵过程中总酸含量的变化Fig.5 Changes of total acid content during fermentation
2.3 鱼露样品中挥发性风味成分分析
由表1可知,处理组1、处理组2、处理组3和舟山商品鱼露分别检测出40、35、32和35种挥发性化合物,其中,处理组1中检出的挥发性化合物种类最多,与处理组2相比,处理组1鱼露样品中醇类、醛类、酯类、呋喃类、含氮含硫化合物相对含量增加,且醛类化合物不仅数量增多,种类也由4种增加至13种,酸类、酮类、烷烃类及其他类的相对含量减少。
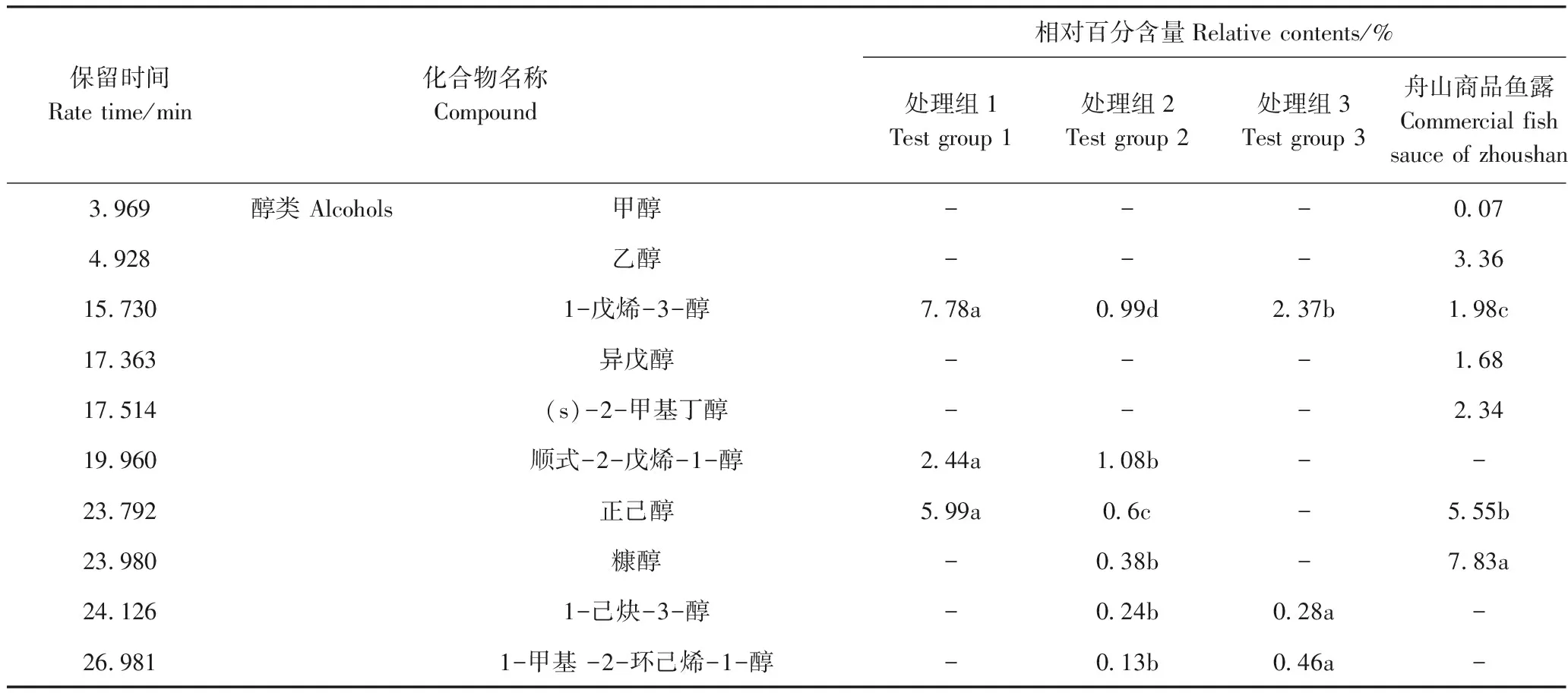
表1 鱼露中挥发性风味物质组成 Table 1 The compositions of volatile compounds in fish sauce

表1(续)
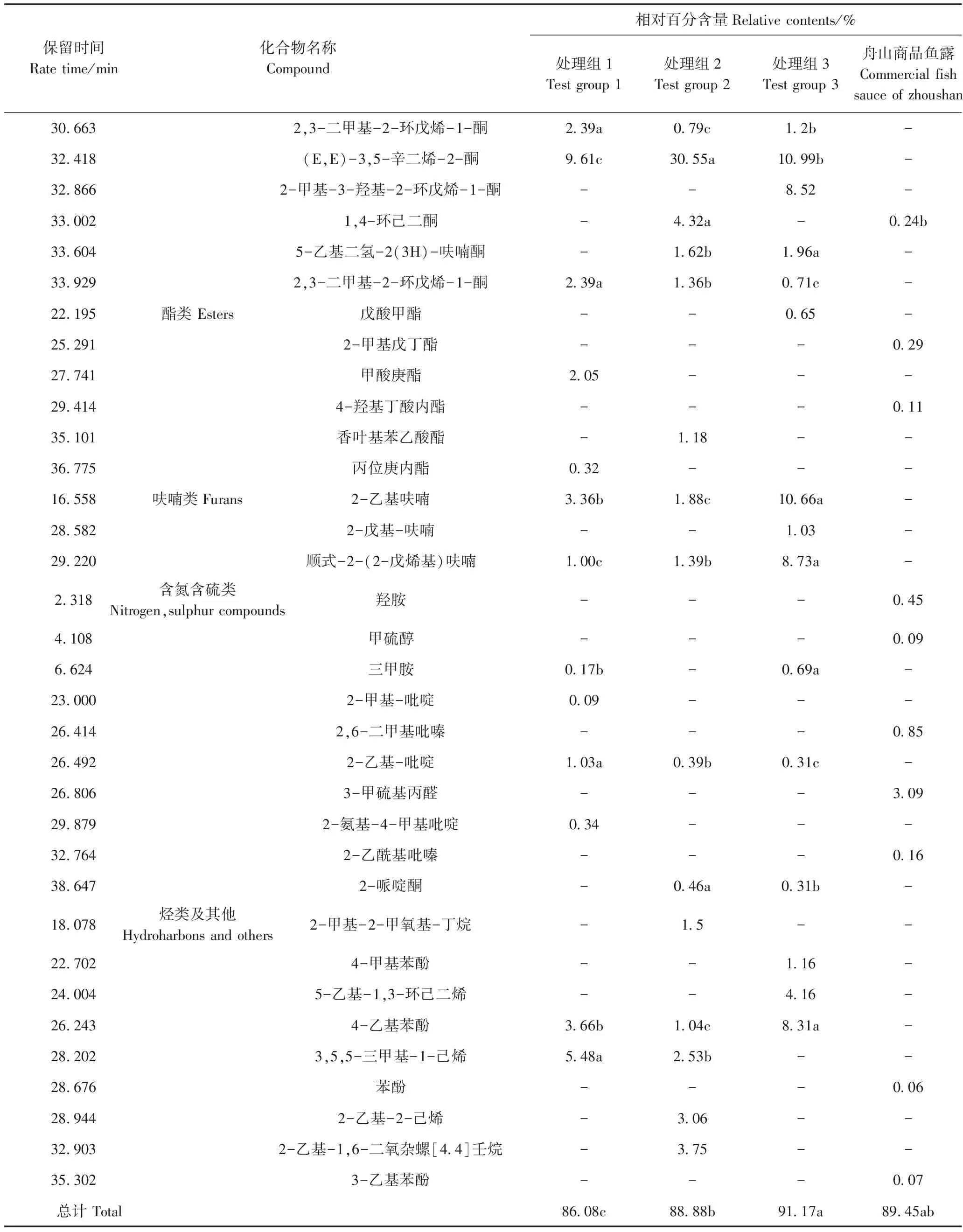
表1(续)
注:“-”表示未检出;相对百分含量=相对峰面积/总峰面积。同行不同小写字母表示差异显著(P<0.05)。
Note:‘-’ means not detected. Relative percentage=relative peak area/total peak area. Different lowercase letters in the peer indicate significant difference at 0.05 level.
醇类化合物大多是由脂肪的氧化分解以及氨基酸和碳水化合物的氧化形成的[22],具有芳香味、植物香、酸败味等。处理组1醇类物质相对百分含量较处理组2、处理组3高,1-戊烯-3-醇是处理组1相对百分含量最高的醇类,达7.78%,该物质能产生烤洋葱香味[23]。低分子量的挥发性酸构成了鱼露的干酪风味[24]。3-甲基丁酸是4种鱼露样品共同检测出的酸类化合物,其中,处理组1酸类化合物的种类和数量低于处理组2,正丁酸是舟山商品鱼露相对百分含量最高的挥发性风味化合物。醛类化合物的阈值比其他化合物阈值低,具有脂肪香味、水果香味和鱼腥味[25]。苯甲醛是4种鱼露样品共同检测出的化合物,能贡献杏仁味[26]。处理组2检测出的(E,E)-3,5-辛二烯-2-酮的相对百分含量高达30.55%,该化合物具有特殊的奶油风味[27],处理组1中(E,E)-3,5-辛二烯-2-酮含量低于处理组2。呋喃类化合物一般认为由脂肪酸的氧化[28]或通过amadori途径重排产生,通常阈值较低,可增强鱼露香气[29],处理组中检测出大量呋喃类化合物。酯类化合物通常在微生物的作用下由醇类和酸类物质经过酯化反应等复杂的生化过程生成[30],处理组1酯类的种类、数量较处理组2、处理组3、舟山商品鱼露均有增加。肖宏艳[31]发现含硫化合物对鱼露风味有重要贡献,处理组1、处理组2、处理组3均未检测出含硫化合物,而舟山商品鱼露中检测出3-甲硫基丙醛,相对百分含量为3.09%,处理组1和处理组3均检出具有鱼腥味的三甲胺。烷烃类及其他化合物在处理组1、处理组2、处理组3、舟山商品鱼露中相对百分含量分别为9.14%、11.88%、13.63%、0.13%。
2.4 鱼露样品感官评价分析
由图6可知,处理组1、处理组2、处理组3及舟山商品鱼露样品的咸味、肉味、鱼腥味差别不明显,其中,处理组1鱼露样品干酪风味和酸味略有不足,但其鲜味、水果香味均优于处理组2和处理3,可能是接种SW5菌株发酵后产生的一些醇类、醛类化合物影响了鱼露的风味。此外,处理组3鱼露样品的腐臭味最大,可能是处理组3未经过巴氏消毒,发酵液中存在大量腐败微生物,即使在高渗透压下仍能生产繁殖,导致处理组3腐臭味较大。

图6 鱼露样品的感官评定雷达图Fig.6 Radar plots depicting the flavor profiles of fish sauce products
3 讨论
本研究表明,贝莱斯芽孢杆菌SW5菌株有2个产酶高峰期,分别在培养26 h和32 h,培养0~26 h时,该菌株分泌的胞外蛋白酶不断在发酵液中积累,并在培养26 h时达到第一个产酶高峰期;至于第二个产酶高峰期,这在贝莱斯芽孢杆菌PS3全基因组(基因登录号为LN999829.1)中可以找到多条胞内蛋白酶基因,正常情况下胞内蛋白酶不会释放至胞外,当细胞进入衰亡期,某些细胞可能产生裂解细胞壁的水解酶,导致胞内蛋白酶释放至胞外[32-33]。AA-N、TVB-N含量是衡量鱼露品质的重要指标,其中,AA-N是以氨基酸形式存在的氮元素含量,该指标越高,说明鱼露中氨基酸含量越高,鲜味越好[34],能反映蛋白质的水解程度及风味特点[35];TVB-N是以氨或胺类等形式存在的碱性含氮物质,是反映原料鱼和肉鲜度的主要指标[36],它的产生是在腐败微生物或酶的作用下,将鱼类加工副产物中的蛋白质或是非蛋白含氮化合物降解为挥发性的碱性胺或是氨。发酵4 d后处理组1发酵醪液AA-N含量增长缓慢,可能是加盐之后高渗透压对该菌株所产蛋白酶活性有抑制作用,导致鱼蛋白的水解速度放缓,发酵1~3 d时TVB-N含量增长较快,可能与其处理方式有关。前期研究可知SW5菌株高产蛋白酶会将鱼蛋白分解为氨基酸。贝莱斯芽孢杆菌PS3全基因组中可以找到多条脱羧酶和脱氢酶基因,鱼露自然发酵前期,鱼蛋白中的某些氨基酸在细菌脱羧酶、脱氢酶的作用下分解为甲胺、二甲胺、三甲胺等碱性含氮物质[37],从而导致TVB-N含量上升。因此,在使用SW5菌株发酵鱼露初期,会造成TVB-N含量上升快,但感官评定结果显示,接种发酵的鱼露无不良风味,可能是由于产生的三甲胺浓度低,不足以影响感官品质。
本研究显示,处理组1鱼露中醇类化合物的相对百分含量高于处理组2和处理组3,与舟山商品鱼露近似,由于醇类化合物的阈值相对较高,所以总体对鱼露的风味贡献不明显,但是可作为风味形成的基础物质[38]。肖宏艳[31]在对潮汕鱼露发酵过程中挥发性风味成分进行分析时发现,风味化合物醇类、醛类、酸类和酮类在鱼露发酵前期均被检出,且醇类化合物含量较高,这与本研究结果较一致。鱼露中的醛类化合物来源于发酵过程中的脂质氧化[39],由甘油二酯在脂肪酶作用下生成的游离脂肪酸进一步氧化形成[1,40]。徐伟芳等[12]报道甲基营养型芽孢杆菌在适宜条件下产生脂肪酶,而接种SW5菌株发酵后醛类化合物的种类、数量均大于未接种发酵和自然发酵,可能是SW5菌株产生的脂肪酶将甘油二酯酶解为游离脂肪酸,之后经过进一步氧化生成了醛类化合物。低分子量的挥发性酸构成了鱼露的干酪风味,处理组1中酸类化合物的种类、相对百分含量较处理组2、处理组3和舟山商品鱼露少,这与pH值、总酸含量和感官评价结果相互印证,处理组1中酸类化合物相对百分含量低可能是鱼露陈酿时间较短,或是由该菌种产酸能力较弱造成的。而处理组2和处理组3的酸类化合物种类数量相当,可能是由于巴氏消毒未完全杀死一些耐热的产酸微生物,在适宜的条件下这些微生物又重新生长繁殖,导致处理组2中含有一定量的酸类化合物。舩津保浩等[41]研究发现,挥发性酸的存在对鱼露的海鲜味有一定的促进作用。Udomsil等[42]使用嗜盐四联球菌(Tetragenococcushalophilus)发酵鱼露时发现,该菌种能提高游离氨基酸和风味物质的含量,又能降低鱼露生物胺含量,说明外接微生物发酵可以有效改善发酵产品的品质,但原料来源不同、接种微生物不同、处理方式不同会使风味结果产生巨大差异,因此结合多种风味指纹图谱是确定风味成分的有效手段[43]。
本研究中,发酵第6天时,处理组1发酵醪液AA-N含量最高,为0.76 g·100 mL-1,表明SW5菌株产蛋白酶能力强,可将鱼蛋白迅速水解,但由于发酵时间短,鱼露样品中挥发性酸类化合物含量较低,且未检测出3-甲硫基丙醛、二甲基二硫、二甲基三硫、丙酸、丁酸等对鱼露挥发性风味有重要贡献的化合物。为改善接种发酵法生产鱼露的风味,后续可采用分段式快速发酵鱼露工艺[21],即前期通过接种贝莱斯芽孢杆菌SW5菌株使鱼蛋白快速水解,再向发酵醪液中接种乳酸菌、产香酵母菌等进行后期发酵,促进鱼露品质成熟,提升鱼露风味[24]。
4 结论
本研究显示,接种贝莱斯芽孢杆菌SW5菌株发酵鳀鱼鱼露,能加速鱼蛋白分解,缩短鱼露发酵生产周期。挥发性风味成分检测结果显示,接种SW5菌株发酵比未接种发酵和自然发酵具有更多种类的挥发性风味物质。因此,贝莱斯芽孢杆菌SW5菌株可用作海洋蛋白质源发酵精深加工的优良微生物菌株,其产蛋白酶的分子机制、酶结构与功能及酶解产物特性等有待进一步研究。
猜你喜欢
分子催化(2022年1期)2022-11-02
食品与发酵工业(2021年23期)2021-12-27
食品与发酵工业(2021年8期)2021-05-10
食品与生活(2020年8期)2020-09-12
皮革制作与环保科技(2020年14期)2020-03-17
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年8期)2019-11-23
当代化工研究(2016年7期)2016-03-20
当代化工研究(2016年5期)2016-03-20
