
铝胁迫对油菜根系形态和生理指标的影响
2019-08-27 06:57韩德鹏刘星月王馨悦付东辉周庆红
核农学报 2019年9期
韩德鹏 刘星月 王馨悦 罗 莎 付东辉 周庆红
(江西农业大学农学院,江西 南昌 330045)
土壤酸化是世界范围内影响农业生产的重要限制因素之一。 近年来,由于环境污染、酸雨沉降及化肥的大量施用导致我国耕地土壤酸化问题进一步加剧[1]。据报道,我国酸性土壤面积约占国土总面积的21%,达到203 万hm2,主要遍及14 个省区,尤以南方分布面积最广[2]。 江西省酸性耕地(pH 值介于4.5 ~5.5之间,平均为5.2)面积为237.48 万hm2,占总耕地面积的84.22%,且酸化程度和面积均呈增加和扩大趋势[3]。 铝(Al)毒是酸性土壤中限制作物生产的主要因素之一,能够影响作物生长,进而导致作物减产[4]。研究发现Al3+是植物受铝毒胁迫的主要形式[5]。 植物对铝毒害的反应首先表现在根系上,铝胁迫1 ~2 h 可观察到植物根系的伸长受到明显抑制[6]。 Kochian等[7]研究表明,铝能够与作物根尖细胞的细胞壁、质膜表面和细胞核等部位结合,从而抑制根尖细胞的伸长与分裂,进而影响作物根系对水分、养分的吸收。 作物根系受到铝胁迫后主根变粗变短,根尖膨大变褐,侧根和根毛减少甚至消失;叶片受到铝胁迫时症状为幼叶变小,叶缘卷曲,叶片变黄,光合作用降低[8-9]。 因此,通过研究铝胁迫对植物根系形态和生理指标的影响,可进一步探讨植物对铝胁迫的生理反应和植物的抗铝机制。
油菜(Brassica napusL.)是我国重要的油料作物,属于十字花科芸薹属。 我国是世界上最大的油菜生产国之一,年均播种面积和总产量均占世界总量的20%以上,我国90%以上油菜栽培于铝毒危害较为严重的长江流域[10]。 研究表明,当土壤pH 值小于5.8,铝浓度大于15 mg·kg-1时油菜就会受到铝毒胁迫,导致产量明显下降[11]。 铝毒危害已经成为限制我国南方油菜产量的重要因素之一。 目前,前人对油菜幼苗耐铝毒的研究多集中在不同油菜品种耐铝毒方面,采用的试验材料较为随机,旨在筛选油菜耐铝相关形态和生理指标鉴定[12-15]。 而关于铝胁迫下油菜根系形态和生理指标的系统性研究尚鲜见报道。 为此,本试验基于公认的耐铝毒指标(主根相对伸长率),前期从300份甘蓝型油菜自交系群体中[16]筛选出的耐铝毒差异显著的2 个品种(R178 和S169),研究不同浓度铝处理对R178 和S169 幼苗根系形态和生理指标的影响,以期为耐铝油菜资源筛选和新品种选育提供理论依据。
1 材料与方法
1.1 试验材料
试验于2018年1-4月在江西农业大学农学院人工气候培养室(光照时间14 h、昼夜25℃/20℃)进行。根据铝胁迫下主根相对生长率和苏木精染色结果,从300 份甘蓝型油菜自交系群体中筛选出耐铝毒差异显著的2 个品种,分别为R178(耐铝油菜品种)、S169(铝敏感油菜品种)作为试验材料,由江西农业大学作物生理生态与遗传育种教育部重点实验室提供。
1.2 试验方法
1.2.1 胁迫处理 取2 个品种的种子,清洗浮选后分别播种在装有纯水的方形塑料盆中纱布上,5 d 后待子叶完全展开、侧根还未从主根分化时,每个品种各选长势一致的60 株幼苗转入装有1/4 营养液(pH 值5.8,华南农业大学叶菜B 配方[17])的方形盆中,培养3 d 后更换成1/2 营养液,再培养3 d 后转入全营养液。 在全营养液中培养10 d 后进行铝胁迫试验,将植株从营养液中取出,转移至0.5 mmol·L-1CaCl2溶液(pH 值4.5)中,处理12 h,分别移入铝浓度为0(对照,CK)、50、100、150、200、300 μmol·L-1的营养液(pH值4.5) 中,每个处理设3 次重复。 铝以氯化铝(AlCl3)形式供给。
1.2.2 幼苗形态指标的测定 铝胁迫处理4 周后,取各材料根系,利用RhizoScan 原位根系扫描仪(加拿大Regent 公司)进行根系扫描,得到根系原位图并存入计算机转换为数字化根系图像后,采用根系图像分析软件WinRHIZO STD4800 LA2 400 分别计算总根长(cm)、根表面积(cm2)、根平均直径(mm)、根体积(cm3)和根尖数(个)。 按照公式计算各材料根系主根相对伸长率:
1.2.3 幼苗生理指标的测定 采用电导率法[18]测定植物细胞膜透性;酸性茚三酮显色法[19]测定游离脯氨酸含量;氮蓝四唑(nitroblue tetrazolium,NBT)法[20]测定超氧化物歧化酶(superoxide dismutase,SOD)活性;愈创木酚法[21]测定过氧化物酶(peroxidase,POD)活性;分光光度法[22]测定过氧化氢酶(catalase,CAT)和抗坏血酸过氧化物酶(ascorbate peroxidase,APX)活性;考马斯亮蓝G-250 染色法[23]测定可溶性蛋白含量。
1.3 数据处理
采用Microsoft Office Excel 2007 整理数据;SPSS Statistics 17.0 进行统计分析。
2 结果与分析
2.1 铝胁迫对油菜幼苗根系形态指标的影响
由图1、图2可知,R178 和S169 的根系形态存在明显差异,同一铝浓度处理下,与S169 相比,R178 在根系发育和地上部生长方面均表现出较强优势。
由表1可知,随着铝处理浓度的增加,2 个品种幼苗的主根相对伸长率均受到明显抑制。 与CK 相比,R178 各处理的主根相对伸长率介于34.3%~95.6%之间,S169 的主根相对伸长率介于20.3%~81.8%之间。 50 μmol·L-1铝处理下,R178 的主根相对伸长率略有降低,但与CK 间未达到显著水平,而S169 与CK相比下降18.2 个百分点;300 μmol·L-1铝处理下,R178 和S169 的主根相对伸长率分别为34.3%、20.3%,说明S169 根系受铝胁迫影响更为严重。
随着铝处理浓度的增加,2 个品种的总根尖数均呈先增加后减少的趋势。 其中,R178 在150 μmol·L-1铝处理时根尖数最多,较CK 增加了31%,而S169 在50 μmol·L-1铝处理时达到最大,较CK 增加了11%。说明在一定铝浓度范围内油菜根尖数随着铝处理浓度的增加而增加,超过某一限度后油菜根尖数不再增加,这可能是低浓度铝处理下主根生长受抑制,油菜总根数的增加,高浓度铝处理下受铝毒害的影响,侧根数增多导致油菜总根数有所减少。 随着铝处理浓度的增加,油菜幼苗的总根长、总根体积、总根表面积和根系平均直径均受到不同程度的影响。 50 μmol·L-1铝处理时R178 的总根长、总根体积、总根表面积和根系平均直径与CK 间均无显著差异,而S169 分别较CK 下降8%、5%、13%、8%。 同一铝浓度处理下,S169 各指标的抑制效果均大于R178,200 μmol·L-1铝处理下,与CK 相比,R178 的总根长、总根体积、总根表面积和根系平均直径降幅分别为24%、19%、26%、18%,S169 的降幅分别为45%、31%、45%、35%。
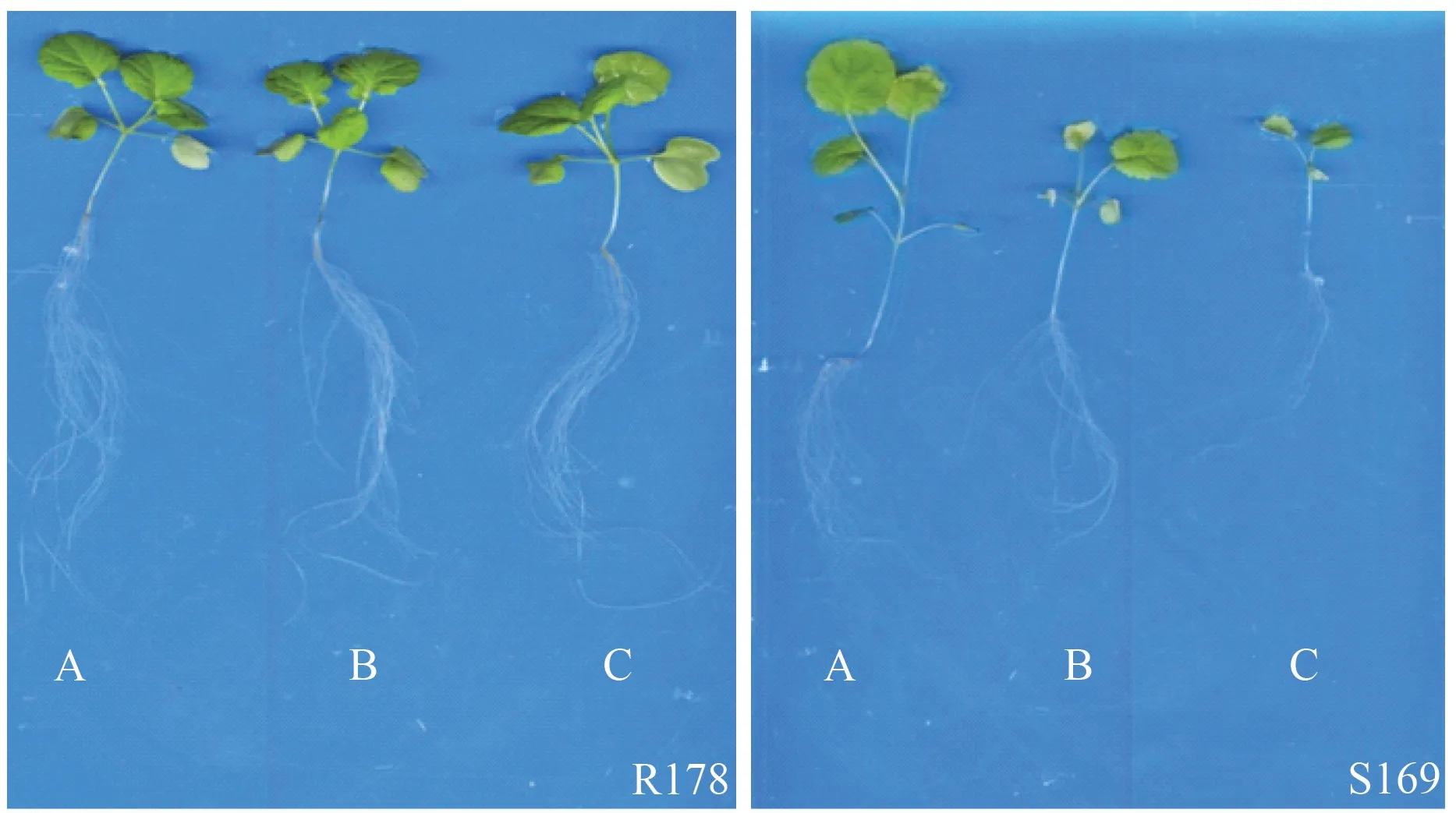
图1 铝胁迫对2 个油菜品种幼苗生长的影响Fig.1 Effect of aluminum stress on seedling growth of two rapeseed varieties
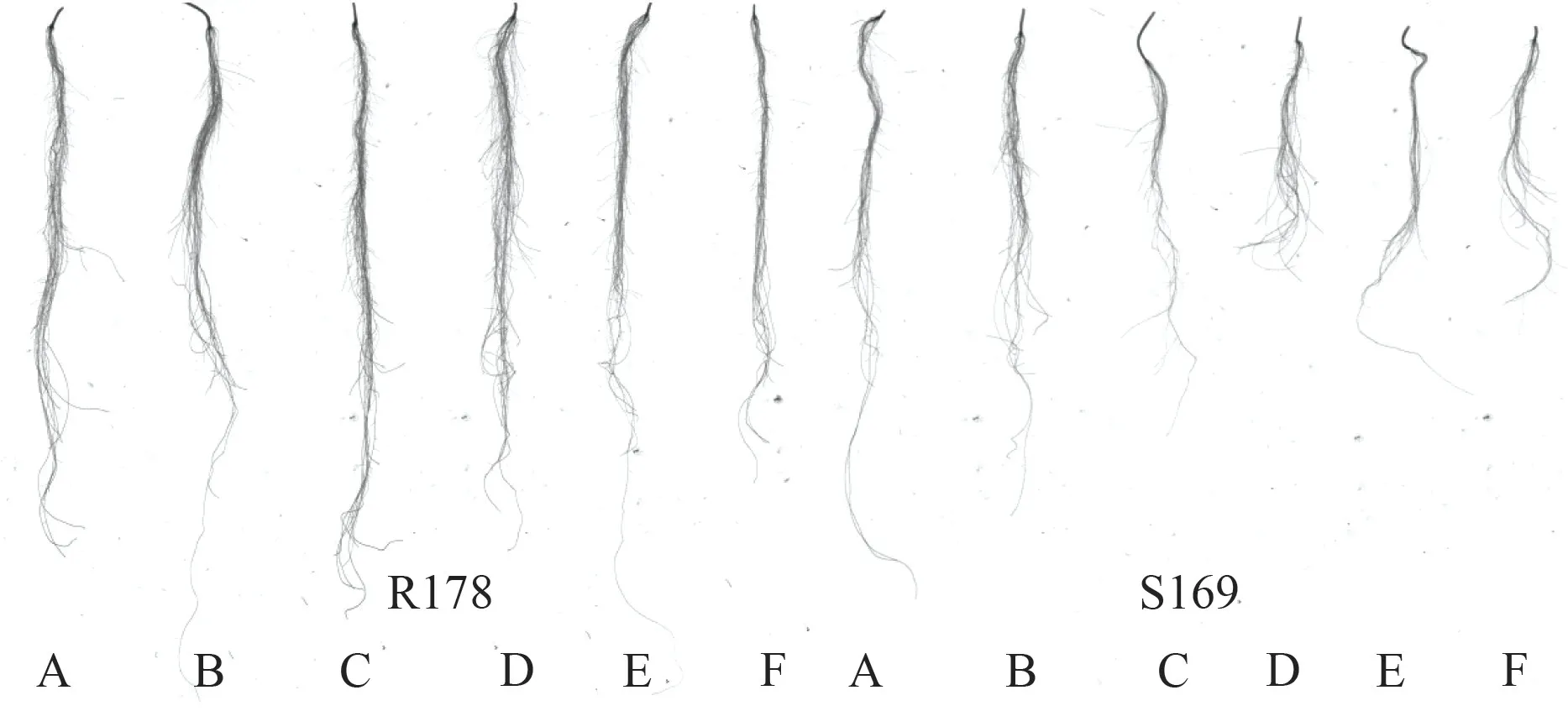
图2 不同浓度铝处理浓度下油菜根系扫描图Fig.2 The scan of rape root with different treatments of aluminum concentrations

表1 铝对油菜根系形态指标的影响Table1 Effect of aluminum on morphological index of root system of Brassica napus L.
2.2 铝胁迫对油菜幼苗根系和叶片生理特性的影响
2.2.1 铝胁迫对油菜幼苗根系和叶片可溶性蛋白质含量的影响 由图3可知,铝处理下2 个油菜品种质根系中的可溶性蛋白质含量均呈先增加后减少的趋势,100 μmol·L-1铝处理时,R178 根系中可溶性蛋白质含量达到最大,为CK 的155%;50 μmol·L-1铝处理时,S169 根系中可溶性蛋白质含量达到最大,为CK 的121%;300 μmol·L-1铝处理时R178 和S169 根系中可溶性蛋白质含量分别降低为CK 的78%和45%,其中S169 根系中可溶性蛋白质积累受到明显抑制。 2 个品种叶片中的可溶性蛋白质含量随着铝处理浓度的增加均呈不断下降的趋势。

图3 铝对油菜根系(A)和叶片(B)可溶性蛋白质含量的影响Fig.3 Effect of aluminum on soluble protein content in root (A) and leaf (B) of Brassica napus L.
2.2.2 铝胁迫对油菜幼苗根系和叶片脯氨酸含量的影响 在生物体内,脯氨酸不仅是理想的渗透调节物质,而且可作为膜和酶的保护物质及自由基清除剂。由图4可知,R178 和S169 根系中脯氨酸含量随着铝处理浓度的增加均呈先增加后减少的趋势,150 μmol·L-1铝处理时二者根系中脯氨酸含量均达到最大,分别为CK 的155%和136%。 2 个品种叶片中脯氨酸含量均呈不断增加的趋势,且均在300 μmol·L-1铝处理时达到最大,其中R178 较CK 增加了一倍,S169 的增幅为66%。 综上表明,油菜根系受铝毒胁迫较叶片严重,无论是根系还是叶片中,R178 的耐铝毒抗性均优于S169。

图4 铝对油菜根系(A)和叶片(B)脯氨酸含量的影响Fig.4 Effect of aluminum on proline content in root (A) and leaf (B) of Brassica napus L.
2.2.3 铝胁迫对油菜幼苗根系和叶片细胞质膜透性的影响 植物细胞膜对维持细胞的微环境和正常的代谢具有重要的作用。 当植物受到逆境影响时,细胞膜遭到破坏,膜透性增大,从而使细胞内的电解质外渗,电导率增大。 由图5可知,铝处理下2 个油菜品种幼苗根系质膜透性均有所增加,各处理下R178 根系相对电解质外渗率较CK 分别增加了2、13、29、47 和54个百分点,除50 μmol·L-1铝处理外,R178 根系相对电解质外渗率与CK 间均差异显著。 S169 根系相对电解质外渗率的变化趋势与R178 相同,200、300 μmol·L-1Al3+处理时2 个品种间均无显著差异,100 μmol·L-1铝处理时R178 和S169 根系相对电解质外渗率分别较CK 显著增加14 和38 个百分点。 50 μmol·L-1铝处理时,R178 的叶片相对电解质外渗率与CK 间无显著差异,而其他铝处理下其相对电解质外渗率均显著高于CK。 300 μmol·L-1铝处理时,S169 叶片的相对电解质外渗率达到最大,但与200 μmol·L-1铝处理时无显著差异。

图5 铝对油菜根系(A)和叶片(B)质膜透性的影响Fig.5 Effect of aluminum on membrane permeability in root (A) and leaf (B) of Brassica napus L.
2.3 铝胁迫对油菜幼苗根系及叶片保护酶活性的影响
2.3.1 铝胁迫对油菜幼苗根系及叶片SOD 活性的影响 由图6可知,R178和S169根系中SOD活性随着铝处理浓度的增加均呈先升高后降低的趋势,各铝处理浓度下R178 根系SOD 活性均高于S169。 R178 叶片中SOD 活性随着铝处理浓度的增加维持在较高水平,但无明显的变化规律;300 μmol·L-1铝处理时,R178 叶片中SOD 活性较CK 显著增加35%;300 μmol·L-1铝处理时,S1690 叶片的SOD 活性显著低于其他铝处理,但仍较CK 显著增加16%。
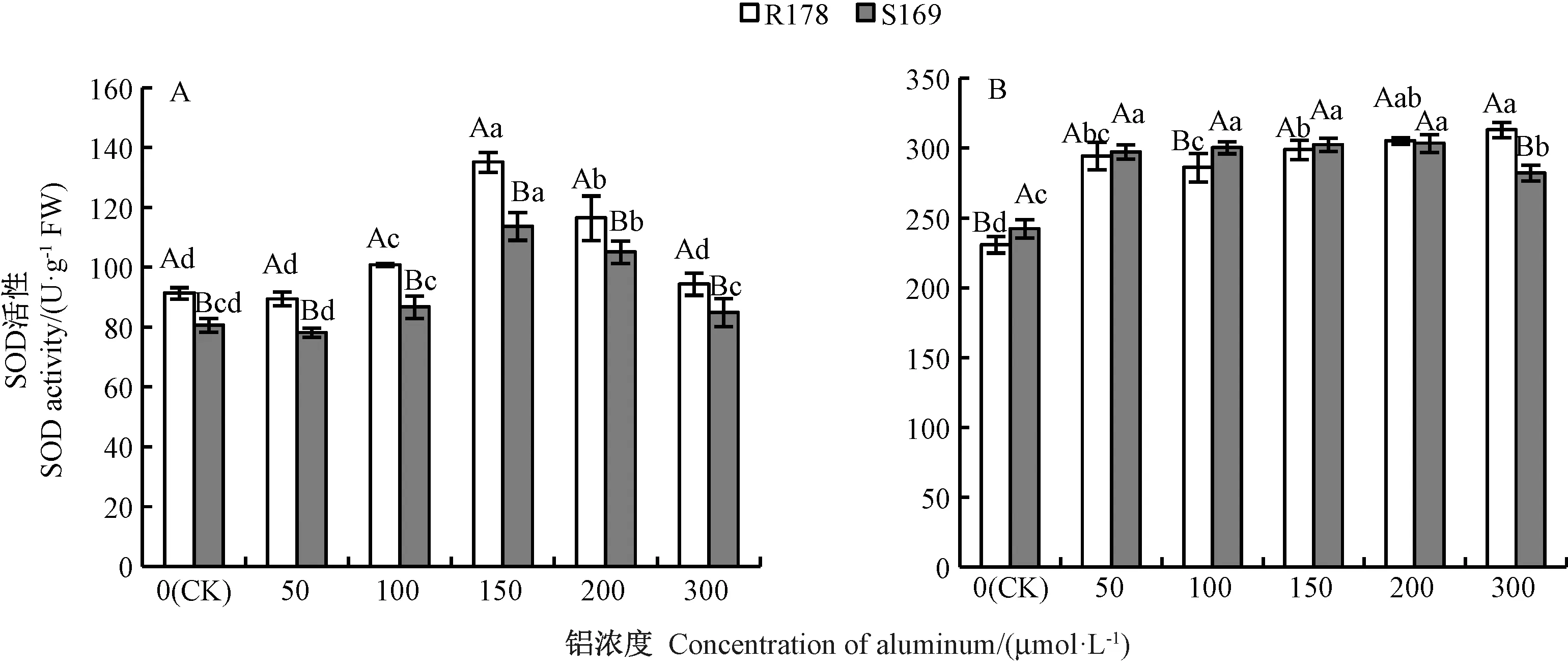
图6 铝对油菜根系(A)和叶片(B)SOD 活性的影响Fig.6 Effect of aluminum on SOD activity in root (A) and leaf (B) of Brassica napus L.
2.3.2 铝胁迫对油菜幼苗根系及叶片POD 活性的影响 由图7可知,铝处理下2 个品种油菜根系中POD活性均远高于叶片,且随着铝处理浓度的增加均呈先增加后降低的趋势。 其中,根系中R178 的POD 活性在100 μmol·L-1铝处理时达到最大,相对CK 的增幅达到22%,与150 μmol·L-1铝处理间无显著差异,但与其他铝处理间均差异显著;100 μmol·L-1铝处理时S169 根系POD 活性最大,但仍显著低于R178。 R178叶片POD 活性在100 μmol·L-1铝处理时达到最大,较CK 高53%,铝处理浓度为150 和200 μmol·L-1时R178 叶片POD 活性无显著差异。 S169 叶片POD 活性在50 μmol·L-1铝处理时达到最大,相对CK 的增幅为40%,且与其他铝处理间均差异显著,铝处理浓度大于50 μmol·L-1时POD 活性显著降低。
2.3.3 铝胁迫对油菜幼苗根系及叶片CAT 活性的影响 由图8可知,铝胁迫下R178 根系的CAT 活性急剧升高,在100 μmol·L-1铝处理时达到最大,较CK 显著增加了86%,当铝胁迫浓度大于100 μmol·L-1时其活性逐渐降低,在300 μmol·L-1铝处理时最小,但与CK 间无显著差异。 S169 根系CAT 活性的变化趋势与R178 相同,除CK 外,同一铝处理浓度下,S169 根系CAT 活性显著低于R178。 R178 叶片中CAT 活性在不同铝处理下较CK 分别显著增加了85%、96%、51%、33%和11%,S169 叶片中CAT 活性较CK 分别增加了34%、64%、25%、19%和5%。
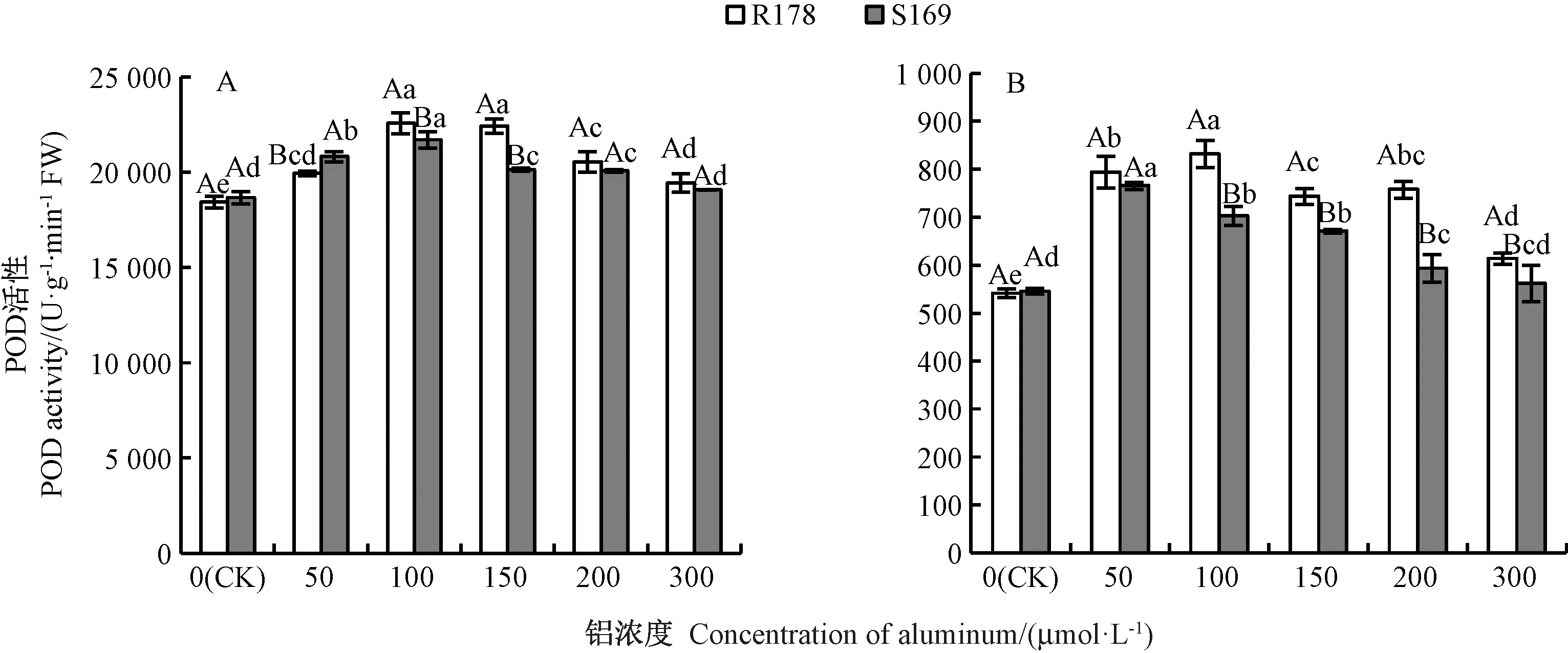
图7 铝对油菜根系(A)和叶片(B)POD 活性的影响Fig.7 Effect of aluminum on POD activity in root (A) and leaf (B) of Brassica napus L.
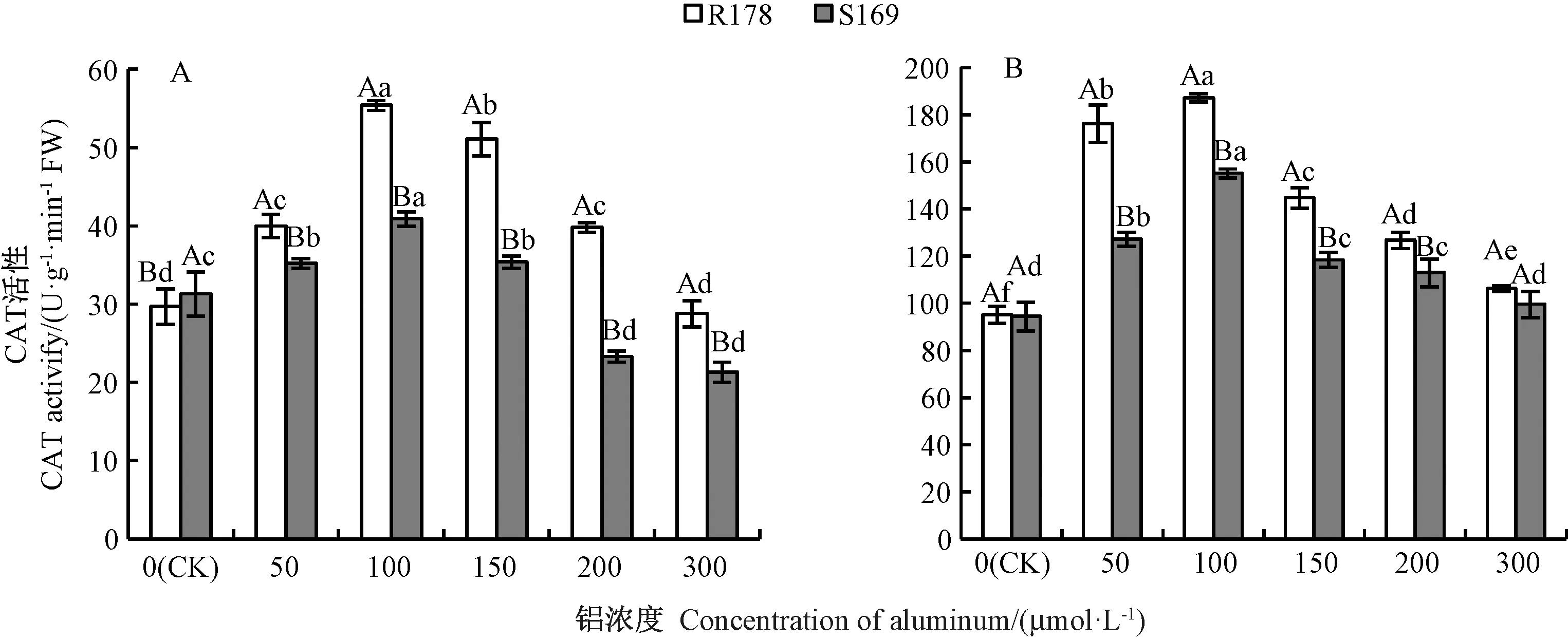
图8 铝对油菜根系(A)和叶片(B)CAT 活性的影响Fig.8 Effect of aluminum on CAT activity in root (A) and leaf (B) of Brassica napus L.
2.3.4 铝胁迫对油菜幼苗根系及叶片APX 活性的影响 由图9可知,除50、200 μmol·L-1铝处理外,相同铝处理下2 个品种根系中APX 活性均无显著差异。50 μmol·L-1铝处理时,R178 叶片的APX 活性与CK间无显著差异,当铝处理浓度高于50 μmol·L-1时,随着铝处理浓度的增加,APX 活性呈先升高再降低趋势,300 μmol·L-1铝处理时较CK 显著降低32%。 S169叶片APX 活性在50 μmol·L-1铝处理时较CK 显著增加16%,100 μmol·L-1铝 处 理 时 达 到 最 大,但 与150 μmol·L-1铝处理间无显著差异,300 μmol·L-1铝处理时较CK 显著降低33%。
3 讨论
研究表明,植物根尖(根冠、分生组织、伸长区)是植物铝积累的主要部位,也是铝毒害的最初作用部位[24]。 铝能够与作物根尖细胞的细胞壁、质膜表面和细胞核等部位结合,从而抑制根尖细胞的伸长与分裂,作物根系受到铝毒害后会导致主根变粗变短,根尖膨大变褐,侧根和根毛减少甚至消失[25-26]。 本研究发现随着铝浓度的增加,油菜幼苗主根相对伸长率显著降低,这与熊洁等[27]和崔雪梅等[28]研究铝毒胁迫下油菜主根相对根长逐渐减小的结论一致。 周楠等[29]研究表明,铝胁迫下,油菜根系形态发生变化,根长和根体积均受到抑制,这与本研究结果基本一致。 本研究中,低浓度铝处理(50 μmol·L-1)对耐铝油菜品种R178 的主根相对伸长率、总根长、总根表面积、根系平均直径和总根体积均无显著影响,但与CK 相比S169的相关指标均受到明显抑制;高浓度铝处理下,2 个品种根系生长均受到显著抑制,且随着铝处理浓度的增加S169 根系生长受抑制程度较R178 明显,说明R178对铝毒抗性显著优于S169。 综上,R178 可作为深入研究油菜耐铝机制的重要材料。
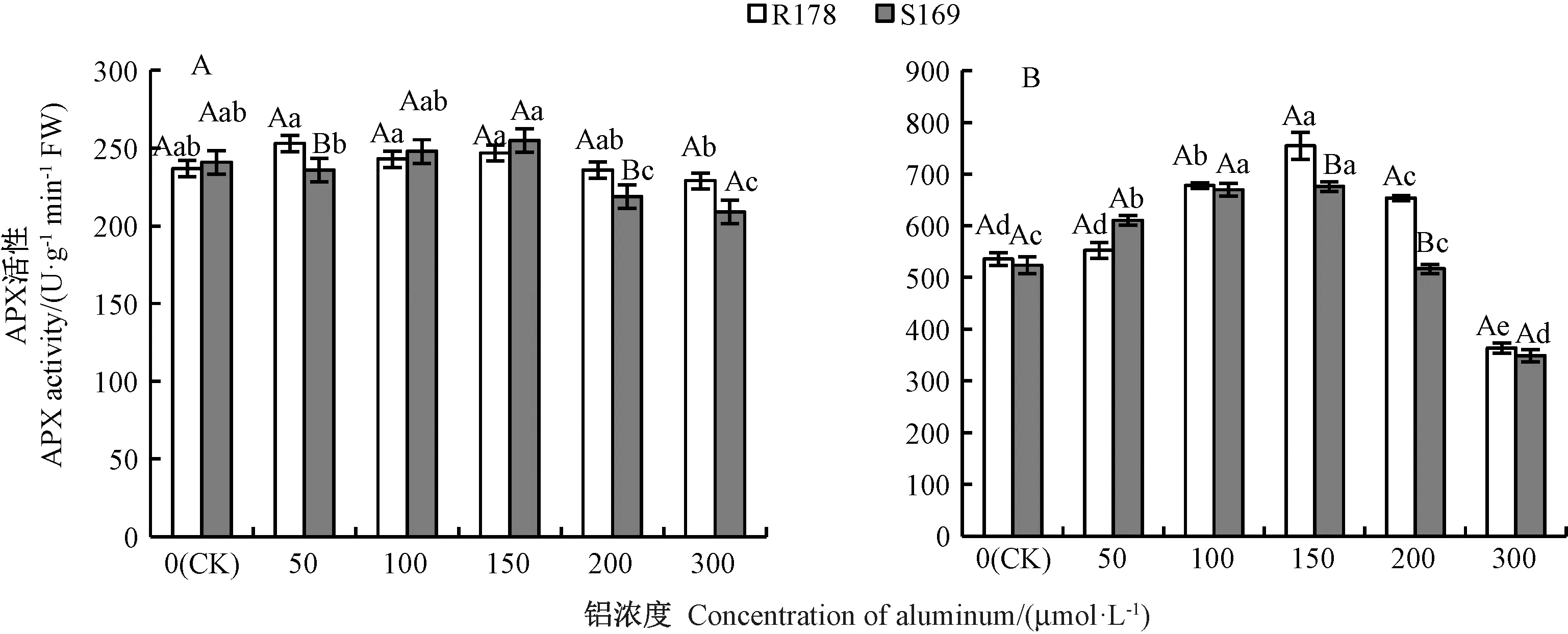
图9 铝对油菜根系(A)和叶片(B)APX 活性的影响Fig.9 Effect of aluminum on APX activity in root (A) and leaf (B) of Brassica napus L.
逆境胁迫下,植物会通过自身代谢变化(如增加脯氨酸含量)抵抗逆境胁迫,脯氨酸可以通过调节及维持细胞渗透压来增加植物的抗逆性[30]。 本研究发现除50 μmmol·L-1铝处理外,S169 根系和叶片中的脯氨酸含量均显著高于CK,2 个油菜品种根系中脯氨酸含量随着铝处理浓度的增加均呈先增加后减少的趋势,叶片中则呈不断增加的趋势;除50 μmmol·L-1铝处理外,2 个品种的叶片中脯胺酸含量无显著差异外,其他铝处理下R178 根系和叶中的脯氨酸含量均显著高于S169,说明R178 可以通过积累较多的脯氨酸来抵抗铝毒害。 可溶性蛋白是重要的渗透调节物质和营养物质,通过提高细胞的保水能力对细胞的生命物质及生物膜起到保护作用,可作为反映植物抗性的指标之一[31-32]。 本研究中,2 个油菜品种根系中的可溶性蛋白质含量随着铝处理浓度的增加均呈先增加后减少的趋势,这可能是由于低浓度铝处理刺激了可溶性蛋白质的快速合成和积累以抵抗不良环境(铝胁迫),高浓度的铝阻碍了蛋白质的合成并导致蛋白质降解。 前人研究发现铝胁迫下,细胞膜结构发生改变、细胞表面离子环境以及跨膜离子通道环境发生改变[33],从而引起胞内的离子、氨基酸、糖类等外渗,质膜透性增加[25]。 本研究中,铝处理浓度低于200 μmol·L-1时R178 根系质膜透性明显低于S169,50 μmol·L-1铝处理时R178 根系和叶片的相对电解质外渗率与CK 间均无显著差异,说明同一铝处理下R178 能够使体内的活性氧含量维持在较低水平,以减少活性氧对质膜的破坏。
SOD 是植物体内天然存在的超氧自由基清除因子,通过歧化反应将有害的超氧自由基转化为H2O2[34];POD 可以清除体内过量的H2O2和过氧化物[35];CAT 是植物细胞内清除H2O2的关键酶,可以将H2O2分解成H2O 和O2[36];APX 是叶绿体内专一清除H2O2的关键酶,在响应逆境胁迫中起到重要作用[37],这些抗氧化酶相互作用,组成一个完整的防氧化系统,可以及时清除植物体内多余的活性氧(reactive oxygen species,ROS),减少细胞膜系统的损伤。 王志颖等[38]研究发现油菜叶片中SOD 活性随着铝浓度的增加呈先减少后增加的趋势,POD 和APX活性均呈增加的趋势,当铝处理浓度为50 μmol·L-1时CAT 活性达到最大值,随着铝浓度继续增加,CAT 活性迅速降低,这与本研究结果不同。 本研究中,2 个油菜品种幼苗根系中SOD、POD、CAT 及叶片中POD、CAT、APX 活性随着铝处理浓度的升高均呈先升高后降低的趋势;100 μmol·L-1铝处理时2 个品系根系和叶中的CAT 活性均达到最大值,随后迅速降低。 造成上述差异的原因可能与供试油菜品种和处理时间有关。 本研究中,低浓度的铝促进了保护酶活性的升高以抵抗逆境,同一处理下耐铝品种R178 的保护酶活性显著高于敏感品种S169,但保护酶清除活性氧的能力是有限的,当铝浓度过高时,植物体内ROS 的产生速率大于清除速率,活性氧无法及时被清除进而导致大量积累,对植物的细胞膜细胞器造成伤害,进而使保护酶活性不断降低。 这与韦东萍等[39]的研究结论相同。 本研究还发现,与油菜根系相比,随着铝浓度的增加油菜叶片中SOD 仍可以维持较高活性,说明叶片受胁迫的伤害较根系轻。
4 结论
铝胁迫对2 个油菜品种幼苗根系生长均有一定抑制作用,耐铝油菜品种R178 通过更多的可溶性蛋白和脯氨酸积累及保护酶活性抵制铝毒胁迫。 本研究结果进一步解析了铝毒胁迫对油菜幼苗生长影响的生理机制,耐铝油菜品种R178 可用于深入开展油菜耐铝分子机制相关研究。
猜你喜欢
中老年保健(2022年3期)2022-11-21
广西植物(2022年5期)2022-06-18
文山学院学报(2022年2期)2022-05-27
课外生活·趣知识(2022年5期)2022-05-14
今日农业(2021年21期)2021-11-26
今日农业(2021年14期)2021-10-14
科学与财富(2019年15期)2019-10-21
发酵科技通讯(2018年2期)2018-07-06
娃娃乐园·综合智能(2018年3期)2018-03-22
数学小灵通(1-2年级)(2017年10期)2017-11-08
