
基于转录组金银花AP2基因家族的生物信息学及表达分析
2019-08-27 06:56乔永刚崔芬芬曹亚萍王勇飞
核农学报 2019年9期
乔永刚 陈 亮 崔芬芬 曹亚萍 王勇飞 宋 芸
(山西农业大学生命科学学院,山西太谷 030801)
低温是限制植物生长和发育的主要环境因子之一。 植物在长期进化过程中形成了一系列复杂而高效的调控网络,以抵御外界低温胁迫[1]。 大量研究表明,植物在抵御逆境胁迫时,包括AP2/ERF、MYB、bHLH、NAC、ZFP、WRKY、VOZ、CAMTA、EIN3 等转录因子起着关键性的调控作用[2-5]。 转录因子是一类和启动子顺式作用元件特异性结合的蛋白,可以与其他蛋白发生相互作用,进而调控下游基因的表达,引起植物对外界环境做出应答反应[2]。 1994年Jofuku 等[6]从拟南芥中成功分离出第1 个AP2 转录因子,进一步分析表明其包含2 个可以与DNA 结合的AP2/ERF 结构域。 不同植物所含有的AP2 转录因子基因家族的数量存在差异,如拟南芥中有147 个,水稻有164 个,玉米167 个,小麦117 个,杨树200 个,葡萄有132 个,而大麦和苹果中分别只有53 和58 个[7-10]。
AP2 转录因子基因家族除参与生物和非生物胁迫过程、植物信号转导途径等,一些AP2 家族成员还是各种逆境信号转导途径中的连接因子[11]。 AP2/ERF家族蛋白主要的结构特征是含有1 个或2 个AP2 结构域,每个结构域约含60 ~70 个氨基酸残基[12]。 根据AP2 家族蛋白结构域中特征元件的种类和数量不同可将其分为3 类,第Ⅰ类含有2 个AP2/ERF 结构域的AP2 家族,在调节植物生长发育过程中起着重要的作用[13-15]。 第Ⅱ类含有1 个AP2/ERF 和1 个B3 结构域的RAV 家族,在植物抵御生物和非生物胁迫过程中发挥着重要作用[16-17]。 第Ⅲ类含有1 个AP2/ERF结构域的ERF 家族,ERF 家族又可以分为2 个亚家族,即CBF/DREB 和ERF 亚家族,CBF/DREB 亚家族可以识别冷诱导响应元件(DRE/CRT),在植物调节抗逆方面起着非常重要的作用[18];而ERF 亚家族与DREB 亚家族的主要区别在于AP2/ERF 结构域的第14 和第19 位氨基酸残基不同,DREB 分别为缬氨酸和谷氨酸,ERF 则分别为丙氨酸和天冬氨酸[19]。
金银花(Lonicera japonicaThunb.)为忍冬科忍冬属植物,以干燥花蕾或带初开的花入药,具有清热解毒、增强人体免疫等功效,是我国重要的中药资源[20]。目前有关金银花AP2 基因家族的研究尚未见报道。本研究对低温胁迫下的金银花进行无参转录组测序,通过Trinity 组装得到相应的Unigenes,挖掘出AP2 转录因子家族基因,对其进行理化性质预测、亚细胞定位预测、磷酸化位点分析、motif 分类并构建系统发育树,以期为金银花AP2 基因家族的功能研究提供一定的理论参考。
1 材料与方法
1.1 试验材料
金银花(品种为大毛花)种苗来源于山东省平邑县,种植于山西省太谷县山西农业大学生命科学学院药用植物研究园,分别取4℃低温胁迫0、3、12 和48 h的叶片为试验材料,样品迅速置于液氮中,并将其送至北京百迈客生物公司,提取总RNA 后构建cDNA 文库,利用Illumina HiSeq 2500 平台进行测序。 对原始测序数据进行过滤、去接头和低质量值数据后,用Trinity 软件进行序列组装并获得Unigene 序列。 从拟南芥转录因子数据库下载AP2 转录因子序列,利用BioEdit 对本地金银花低温转录组数据进行BLAST 比对,筛选出金银花AP2 基因并利用SMART 和Pfam 验证,剔除不完整的基因[21]。
1.2 金银花AP2 基因家族的生物信息学分析
对筛选出的AP2 转录因子的氨基酸序列,利用在线Expasy 生物信息学工具对其进行理化性质分析;利用SubLoc v1.0 进行亚细胞定位分析;利用NetPhos 在线工具进行潜在的磷酸化位点预测分析。 利用MEME在线工具对34 个AP2 基因家族蛋白进行motif 预测;利用MEGA7 软件对34 个蛋白序列进行多序列比对并分类并构建系统发育树,每个计算点计算1 000 次。
1.3 RNA 提取、cDNA 合成及实时荧光定量PCR 验证
按照植物RNA 提取试剂盒(北京华越洋生物技术有限公司)说明书提取金银花根、茎和叶样品的RNA 并反转录成cDNA,按照SYBRPremix Ex TaqTM(Tli RNaseH Plus)试剂盒(北京全式金生物技术有限公司)说明书进行实时荧光定量PCR,扩增程序详见表1;每个样品均设3 次生物学重复,以山西农业大学生命科学学院药用植物遗传育种实验室筛选的ACT2/7 和c49672.graph_c0 为内参基因进行实时荧光定量PCR,采用2-△△Ct法[22-23]计算基因的相对表达量。 所用引物详见表2。

表1 RT-qPCR 反应程序Table1 RT-qPCR protocol

表2 候选内参基因引物信息Table2 Information of candidate reference gene
1.4 数据统计与分析
利用SPSS 20.0 进行数据统计与分析,采用Origin 8.0 软件绘图。
2 结果与分析
2.1 金银花AP2 基因家族理化性质分析
对筛选出34 个具有完整AP2 结构的转录因子,利用Expasy 对AP2 基因家族编码的氨基酸进行理化性质分析。 由表3可知,34 个AP2 基因编码蛋白的氨基酸数量为74 ~887 个;pI 大小为4.74 ~10.54,表明不同的AP2 蛋白在不同的微环境中的生物学功能存在差异;除AP2_10 为稳定蛋白外,其余均属于不稳定蛋白(不稳定指数小于40 为稳定蛋白,大于40 为不稳定蛋白);34 个AP2 蛋白的脂肪系数为42.30~98.03;34 个AP2 基因编码的蛋白的平均亲疏水性均小于0,均属于亲水性蛋白。
2.2 金银花AP2 基因家族亚细胞定位预测
利用SubLoc v1.0 对金银花AP2 基因家族进行亚细胞定位预测,结果显示金银花AP2 基因家族蛋白除AP2_24 和AP2_28 定位于叶绿体外,其余主要定位于细胞核,这符合其作为转录因子调控下游基因表达的特性[24]。 然而AP2_15、AP2_17、AP2_30 和AP2_33在线粒体中也有分布,AP2_34 在叶绿体中同样有分布,说明不同AP2 家族成员在逆境环境下于不同细胞器中发挥着不同的作用。
2.3 金银花AP2 蛋白磷酸化位点分析
蛋白的磷酸化在植物信号转导过程中起着关键作用[25]。 利用NetPhos 对34 个金银花AP2 蛋白的磷酸化位点分析,由表4可知,除AP2_1、AP2_5、AP2_7、AP2_14 和AP2_28 没有酪氨酸(Try)磷酸化位点外,其他金银花AP2 蛋白均具有丝氨酸(Ser)、苏氨酸(Thr)和酪氨酸(Try)磷酸化位点,且除AP2_16 和AP2_31 外,磷酸化位点数依次为丝氨酸>苏氨酸>酪氨酸。
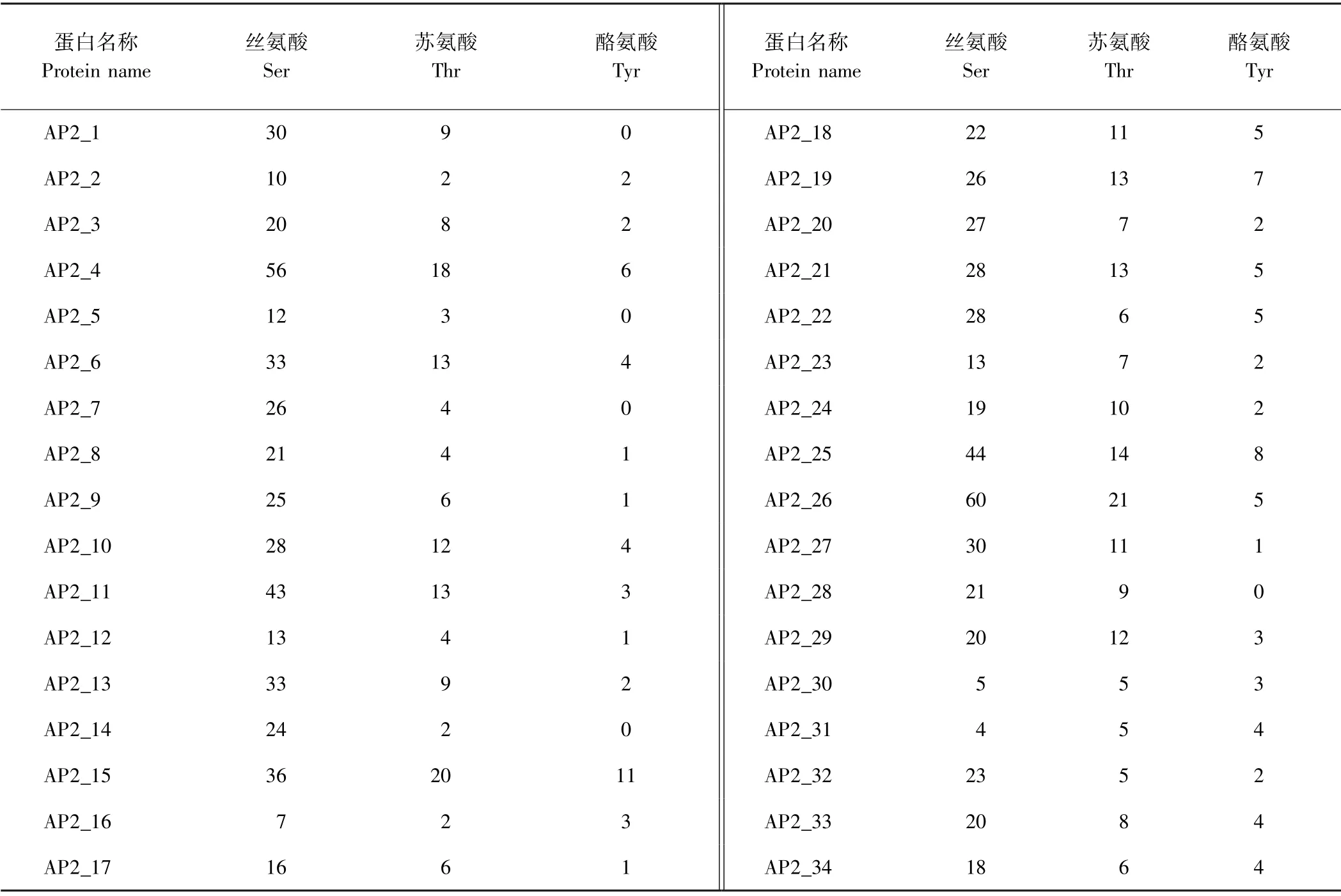
表4 AP2 蛋白的磷酸化位点预测Table4 Phosphorylation site prediction of AP2 protein
2.4 金银花AP2 基因家族蛋白基序及分类
利用MEME 数据库对金银花AP2 基因家族蛋白进行序列分析,结果显示,34 个金银花AP2 基因含有不同数目的保守motif 结构,AP2 家族蛋白包含2 个保守的结构域,RAYD 和YRG,与前人研究一致[26]。motif 1 为RAYD,motif 2 为YRG,而motif 3 则同时包含RAYD 和YRG。 每个AP2 基因家族的成员都含有数量不等的RAYD 和YRG,且不同的AP2 基因家族之间的保守结构域的氨基酸存在变异(图1、图2)。
2.5 金银花AP2 家族的同源性分析
运用MEGA7 软件对筛选的34 个金银花AP2 转录因子和拟南芥的AP2、ERF 以及RAV 转录因子进行序列同源比对,并构建系统进化树。 由图3可知,金银花AP2_10 与拟南芥RAV 家族聚为一类,为RAV 家族;AP2_7、AP2_24 和AP2_30 分别与DREB 的A5、A1和A4 亚族聚为一类;AP2_4、AP2_15、AP2_16、AP2_17、AP2_18、AP2_21、AP2_25、AP2_27、AP2_33 和AP2_34 聚为一类为AP2 家族成员;AP2_12、AP2_16、AP2_6 和AP2_11 与ERF 的B6 亚族聚为一类,AP2_20 与ERF 的B4 亚族聚为一类,AP2_23、AP2_3、AP2_8 和AP2_5 与ERF 的B1 亚族聚为一类,AP2_2、AP2_9、AP2_13、AP2_14、AP2_21、AP2_22、AP2_28、AP2_29、AP2_31 和AP2_32 与拟南芥ERF 的B3 亚族聚为一类;AP2_26 单独聚为一枝,推测其可能是金银花中特有的转录因子。
2.6 DREB 和RAV 基因家族在低温胁迫下的表达分析

图1 AP2 蛋白的基序Fig.1 The motif of AP2 protein

图2 AP2 结构域的基序Fig.2 The motif of AP2 domain
分析聚类结果检测到的3 个DREB(AP2_7、AP2_24、AP2_30)基因和1 个RAV(AP2_10)基因在4℃低温下处理不同时间的表达情况。 由图4可知,不同低温处理时间下的不同组织中的表达水平存在明显差异,DREB基因家族在叶中的表达量明显高于根和茎,且均在低温处理3 h 后表达量达到最高值。 随着低温胁迫时间的增加,DREB基因在根、茎、叶中的表达量整体上均呈先升高后降低再升高的趋势。 而RAV基因在低温处理3 h 后在根中的表达量极显著高于其他处理,且明显高于茎和叶中的表达量,随着低温胁迫时间的增加,根、茎、叶中的表达量整体均呈先升高后降低的趋势。 综上,转录因子在低温胁迫初期开始发挥作用,可能对低温的响应起着关键的作用。
3 讨论
研究表明,AP2 转录因子广泛存在于植物基因组中,调控植物器官的生长发育[27-28]、胁迫响应[29-31],以及药用植物活性成分的生物合成[32-33]等。 目前,关于金银花AP2 转录因子家族的功能研究尚未报道,随着二代测序技术的发展,利用转录组测序数据分析金银花AP2 基因家族已成为可能。
本研究从金银花中筛选了34 个AP2 转录因子,数量明显高于葡萄(20 个)[34],但远低于水稻(164)[8]、拟南芥(147)[8]和杨柳(173 个)[35]等,表明AP2 转录因子在不同的物种进化过程中存在一定的扩张或收缩。 蛋白基序分析发现,不同类型的AP2 转录因子包含的保守基序存在差异,但均包含YRG 和RAYD 两个保守结构域,这与拟南芥[36]、水稻[37]AP2转录因子家族的研究结果一致,表明AP2 转录因子家族在物种进化过程中高度保守,但保守的氨基酸中存在较大的变异,可能是植物在进化过程中为了适应环境而产生的。
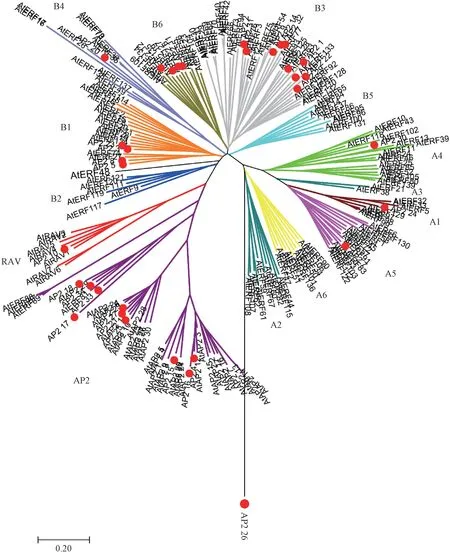
图3 金银花AP2 家族与拟南芥蛋白序列进化树分析Fig.3 Phylogenetic analysis of AP2 family in Lonicera japonica Thunb.and Arabidopsis thaliana protein sequences
同源比对及系统进化发育分析发现,金银花中的AP2_7、AP2_24 和AP2_30 与拟南芥的DREB A5、DREB A1、DREB A4 亲缘关系较近。 在拟南芥中发现ERF105(属于DREB A4)能够响应低温胁迫[38]。 通过RT-qPCR 发现,金银花的AP2_7、AP2_24 和AP2_30在低温胁迫下高表达,且叶中的表达量明显高于茎和根,推测其可能介导叶的抗低温表达。 金银花AP2_10转录因子与拟南芥RAV1 亲缘关系较近。 已有研究表明,拟南芥RAV1 能有直接调控ABI基因,进而在ABA的信号转导过程中发挥重要功能[39]。 RT-qPCR 结果显示,金银花AP2_10 在低温胁迫下3 h 时表达量显著增加,且在根中的表达水平明显高于茎和叶,推测其可能介导了根的抗低温表达。 本研究还发现,金银花AP2_26 未与拟南芥的任何一类聚为一簇,推测其可能是金银花中特有的一类AP2 转录因子,其功能有待进一步研究。 表明,同一亚家族的金银花的AP2 基因具有相对一致的表达模式,系统进化关系相近的蛋白结构相对保守且存在差异,但其所发挥的生物学功能基本相似。

图4 低温胁迫下金银花AP2 基因的表达水平Fig.4 Expression level of AP2 gene in Lonicera japonica Thunb.under low temperature stress
本研究从金银花中筛选出34 个AP2 转录因子,理化性质研究结果表明,不同的AP2 转录因子编码的蛋白的理化性质存在一定的差异,说明其发挥的生物学作用也不相同。 亚细胞预测结果显示,基本所有的AP2 转录因子均定位在细胞核中,这符合其作为转录因子调控下游基因表达的特性,但AP2_24 和AP2_28定位于叶绿体中,其具体功能还有待进一步研究;磷酸化位点分析发现,不同的AP2 的磷酸化位点的个数以及类型存在较大的差异,说明不同的AP2 在植物面对非生物逆境胁迫时,参与了不同的信号转导途径;对金银花AP2 转录因子的保守基序分析显示,所有的AP2均含有其保守的功能结构域YRG 和RAYD,但其保守的氨基酸中存在较大的变异,其原因可能是植物在进化过程中为了适应环境胁迫;对所有的AP2 转录因子进行系统发育分析,发现其与许多模式植物一样含有AP2 家族的4 个亚家族,而DREB主要参与低温逆境胁迫响应,RAV在低温、高温和干旱条件下诱导表达,因此,对筛选出的DREB和RAV家族的基因进行实时荧光定量PCR 表达分析,发现在低温胁迫初期,这两类转录因子就开始大量表达,其在不同的组织器官中表达量存在差异,DERB集中在叶中表达,RAV集中在根中表达,但其表达模式基本相同,推测其可能参与了金银花的低温响应。 综上所述,同一亚家族的AP2 基因具有相对一致的时空表达模式,系统进化关系相近的蛋白结构相对保守且存在差异,但其发挥的生物学功能相似。
4 结论
本研究利用金银花转录组数据共筛选了34 个AP2 转录因子。 系统进化发育分析表明,金银花AP2转录因子家族未能包含拟南芥所有类型的AP2,同时也存在1 个AP2(AP2_26)未能与拟南芥中的AP2 聚成一簇,表明金银花AP2 转录因子家族在进化过程中存在基因的扩张和缺失现象。 4 个AP2 基因在根、茎和叶中的表达分析结果显示,AP2_7、AP2_24 和AP2_30 在叶中高表达,AP2_10 在茎中高表达,但其均可能受低温诱导表达。 本研究结果为进一步研究金银花AP2 转录因子的功能奠定了理论基础。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17
今日农业(2021年15期)2021-11-26
小学生作文(低年级适用)(2019年12期)2020-01-18
广州大学学报(自然科学版)(2019年1期)2019-05-07
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
公民与法治(2016年11期)2016-05-17
华人时刊(2016年13期)2016-04-05
天津科技大学学报(2016年1期)2016-02-28
