
反式-4-羟基-L-脯氨酸的研究进展
2018-07-06 11:38,,,,,2,,2,,2
发酵科技通讯 2018年2期
, , ,,,2,,2,,2
(1.浙江工业大学 绿色制药协同创新中心,浙江 杭州 310014;2.浙江省长三角生物医药产业技术研究园,浙江 德清 313200)
L-脯氨酸(L-proline)是一种结构上带有五元环的天然氨基酸,其五元环上的3,4,5位都可以被羟基化形成羟基脯氨酸,每个位置的羟基化又包含了顺式和反式两种结构,相应的共有6种不同构型的脯氨酸衍生物,包括顺式-3-羟基-L-脯氨酸、反式-3-羟基-L-脯氨酸、顺式-4-羟基-L-脯氨酸、反式-4-羟基-L-脯氨酸、顺式-5-羟基-L-脯氨酸和反式-5-羟基-L-脯氨酸.其中,反式-4-羟基-L-脯氨酸是天然存在于动物胶原蛋白中的一种L-脯氨酸羟基化衍生物[1],在几种羟基化产物中存在最广泛,因此通常将反式-4-羟基-L-脯氨酸简称为羟基脯氨酸,笔者将对羟基脯氨酸的性质、功能、用途和生产方式加以详细论述.
1 羟基脯氨酸的性质
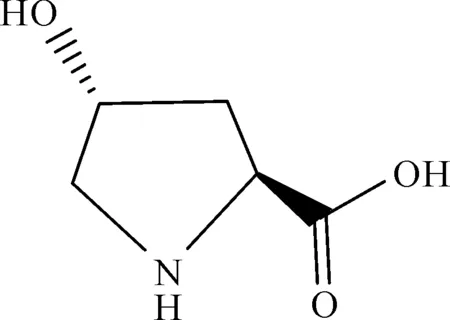
反式-4-羟基-L-脯氨酸,呈白色片状晶体或粉末,分子式C5H9NO3,分子量为131.10,常温常压下较稳定,极微溶于乙醇,不溶于乙醚,25 ℃时在水中的溶解度为361.1 g/L.羟基脯氨酸的化学结构式为
2 羟基脯氨酸的代谢、功能和应用
2.1 生物体内羟基脯氨酸的代谢
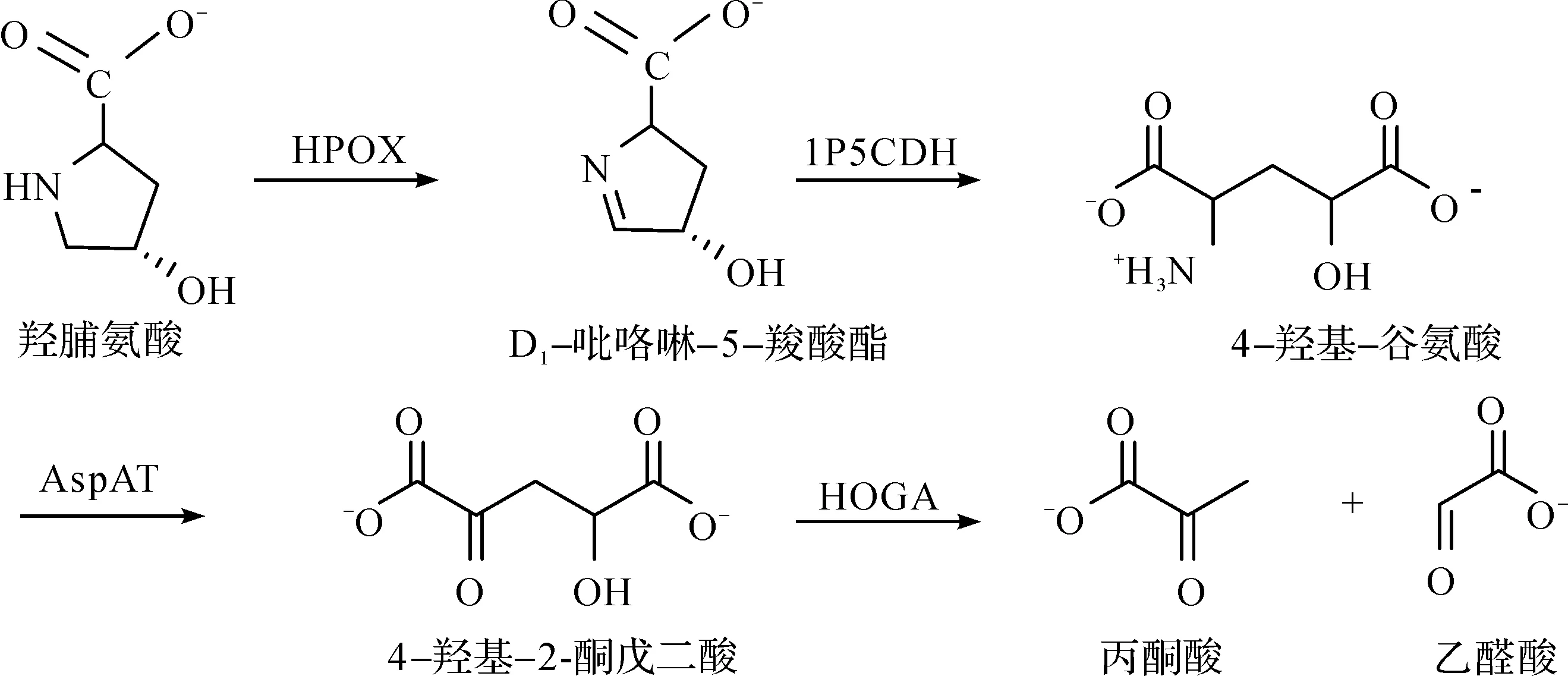
生物体中胶原蛋白的脯氨酸残基在氧气、抗坏血酸、α-酮戊二酸和亚铁离子存在的情况下,可被脯氨酰4-羟化酶催化生成相应的羟基脯氨酸[2],生物体内游离的4-羟基脯氨酸由胶原蛋白降解产生[3].人体内源性胶原蛋白每天能产生300~450 mg的羟基脯氨酸,另外的羟基脯氨酸来源于饮食[4].大部分羟基脯氨酸在肝脏和肾脏中代谢,少于30 mg的羟基脯氨酸通过尿液排出体外[5].大型哺乳动物中羟基脯氨酸的分解代谢途径涉及4种线粒体酶[6]:羟脯氨酸氧化酶(HPOX)、D1-吡咯啉-5-羧酸酯脱氢酶(1P5CDH)、谷氨酸-天冬氨酸转氨酶(AspAT)和4-羟基-2-酮戊二酸醛缩酶(HOGA).前3种酶负责将羟基脯氨酸环氧化成D1-吡咯啉-5-羧酸酯、自发开环和进一步氧化生成4-羟基-谷氨酸,并将4-羟基-谷氨酸转化为4-羟基-2-酮戊二酸(HOG),在4-羟基-2-酮戊二酸醛缩酶催化下,HOG被分解产生丙酮酸和乙醛酸[7].羟脯氨酸的代谢过程为
2.2 羟基脯氨酸的生理功能
羟基脯氨酸是一种非必需氨基酸,主要存在于动物的胶原蛋白中[1],其作用是加强结缔组织的弹性和韧性[2].胶原蛋白的螺旋区域是由多个Gly-X-Y的重复序列组成的,其中脯氨酸可以在X或Y位置,而羟脯氨酸仅存在于Y的位置上[8],其羟基有助于胶原蛋白三螺旋结构的稳定[9].脯氨酸的羟基化反应也发生在其他蛋白质中以调节细胞对缺氧的生理反应[10].近年来的研究发现:羟基脯氨酸在维持植物、动物和人类细胞的细胞结构和功能方面起着重要的作用.比如,在动物体内羟基脯氨酸代谢生成的乙醛酸可以在丙氨酸乙醛酸转氨酶的催化下生成鸡所必需的甘氨酸[11].羟基脯氨酸是细胞代谢的重要调节剂,能够提高体内营养物质利用效率和改善生物体的健康,当一些氨基酸前体的常用来源不可利用时,羟脯氨酸可以在微环境中为氨基酸代谢途径提供所需要的内源性营养[3].
2.3 羟基脯氨酸的应用
由于特有的化学结构和生理功能,羟基脯氨酸已经广泛应用于医药、化工、美容和疾病诊断等方面.
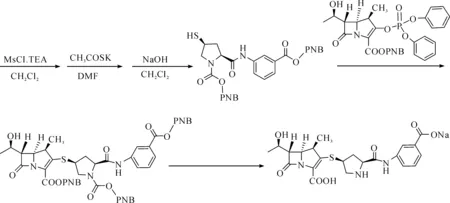
在医药方面,羟基脯氨酸作为起始原料,可以用来合成多种碳青霉烯类抗生素[12],如美罗培南、厄他培南等,此类抗生素具有抗菌谱广、抗菌活性强、毒性及耐药性低、对β-内酰胺酶稳定等特点,对于某些头孢类抗生素无法治愈的感染,培南类药物具有良好的效果[13].郑子新等发现以羟脯氨酸为重要组份研制的注射液对急慢性肝病所导致的低蛋白血症有一定的疗效[14].羟基脯氨酸合成厄他培南的路径为

在手性合成方面,羟基脯氨酸作为手性化合物可以经催化合成多聚物[15],用来分离手性物质.2015年,研究人员将反式-4-羟基-L-脯氨酸键合手性配体制作成色谱固定相,在高效液相色谱中显示可以拆分7种手性化合物、氯酚及二硝基苯两种位置异构体[16].
在美容方面,羟脯氨酸具有消除氧化剂和调整细胞的氧化还原状态的潜在效果[17],有人将羟脯氨酸或N-酰化羟脯氨酸衍生物用于一些高端护肤品中[18].
在疾病诊断方面,羟脯氨酸是结缔组织中唯一特征氨基酸,在胶原降解期间,尿和血清中释放羟脯氨酸的含量与纤维化呈现相关性,在肝疾病诊断中可将羟脯氨酸含量用作纤维化评分的诊断标志[19].此外研究还发现肌肉骨骼系统损害与尿中羟脯氨酸排泄量有一定关系[20],在体内检测尿液中羟脯氨酸的量是判断组织中胶原蛋白损伤的一个生化指标[21].
3 羟基脯氨酸的生产方法
羟基脯氨酸的生产方法主要有动物组织提取法、全细胞催化法和直接发酵法.
3.1 动物组织提取法制备羟基脯氨酸
提取法主要以动物胶原蛋白为原料,经过强酸水解、亚硝酸氧化和离子交换等过程获得羟基脯氨酸.李娟等以明胶为原料,用盐酸水解,亚硝酸钠氧化,D61大孔径阳离子交换树脂分离提取,浓缩结晶后羟基脯氨酸的提取率平均值达到41.4%[22],水解法提取羟基脯氨酸的工艺简单、技术门槛低,但强酸水解胶原蛋白容易造成氨基酸的消旋变性,影响产物的品质.同时废水排放多,环保压力大,目前已基本上被淘汰,其工艺路线为
3.2 全细胞催化法
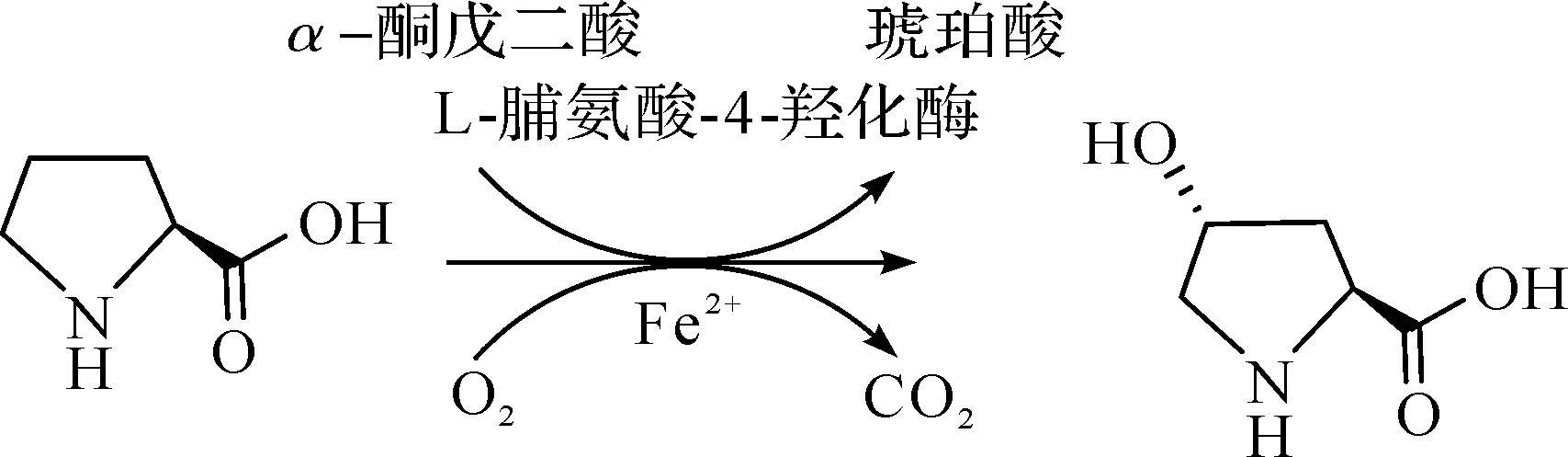
L-脯氨酸-4-羟化酶能够以L-脯氨酸、α-酮戊二酸和氧气为底物,催化生成羟基脯氨酸、琥珀酸和二氧化碳.全细胞催化法和直接发酵法都是以该酶为基础建立起来的.通常将需要添加L-脯氨酸为底物生产羟基脯氨酸的方法定义为全细胞催化法,将以葡萄糖等初级碳源直接发酵生产羟基脯氨酸的方法定义为直接发酵法.
1994年研究者通过对灰色链霉菌(Streptomycesgriseoviridus)的L-脯氨酸-4-羟化酶对α-酮戊二酸依赖性的特异性研究,首次使生物法生产羟基脯氨酸成为可能[23].但是由于酶的不稳定性及难于提取分离等特点,一直限制着体外羟基脯氨酸的合成.
20世纪末,Shibasaki等采用高效灵敏的筛选系统,筛选出具有脯氨酸-4-羟化酶活性的指孢囊菌RH1(Dactylosporangiumsp.),从该菌株中分离纯化得到到反式-4-脯氨酸羟化酶,该酶能够在体外催化L-脯氨酸羟基化生成游离的反式-4-羟基-L-脯氨酸,其特点是具有高度的区域和立体特异性.研究人员将表达反式-4-脯氨酸羟化酶的基因克隆并在大肠杆菌中表达,大肠杆菌重组体显示脯氨酸-4-羟化酶活性,比原始菌株指孢囊菌中的活性高13.6倍[24].脯氨酸-4-羟化酶催化过程为
由于α-酮戊二酸价格较高,因此直接以L-脯氨酸和α-酮戊二酸为底物,催化生成羟基脯氨酸的方案成本过高.所以全细胞催化合成羟基脯氨酸一般只添加外源的L-脯氨酸,而α-酮戊二酸是通过培养过程中TCA循环来供应.Shibasaki等将L-脯氨酸羟化酶在大肠杆菌W1485中表达,在含有L-脯氨酸和葡萄糖的培养基中培养100 h产生并累积41 g/L的羟基脯氨酸,使L-脯氨酸到羟基脯氨酸的转化率为87%.在此基础上,该课题组进一步敲除了大肠杆菌W1485菌株中的putA基因,阻断了L-脯氨酸到谷氨酸的代谢途径,虽然同样条件下羟基脯氨酸的最终产量还是41 g/L,但L-脯氨酸的转化率从原来的87%提高到100%[25].此外,全细胞催化法合成羟基脯氨酸的研究主要从反式-4-脯氨酸羟化酶基因序列入手,通过添加分子伴侣、优化反式-4-脯氨酸羟化酶基因序列等[26-27]来提高反式-4-脯氨酸羟化酶的酶活.例如,刘合栋等对脯氨酸-4-羟化酶基因优化与强启动子串联得到高表达的脯氨酸羟化酶[28],经过添加外源L-脯氨酸和发酵条件的优化,羟基脯氨酸的产量达到了42.5 g/L[29].表1是不同全细胞催化法合成羟基脯氨酸的总结对比.
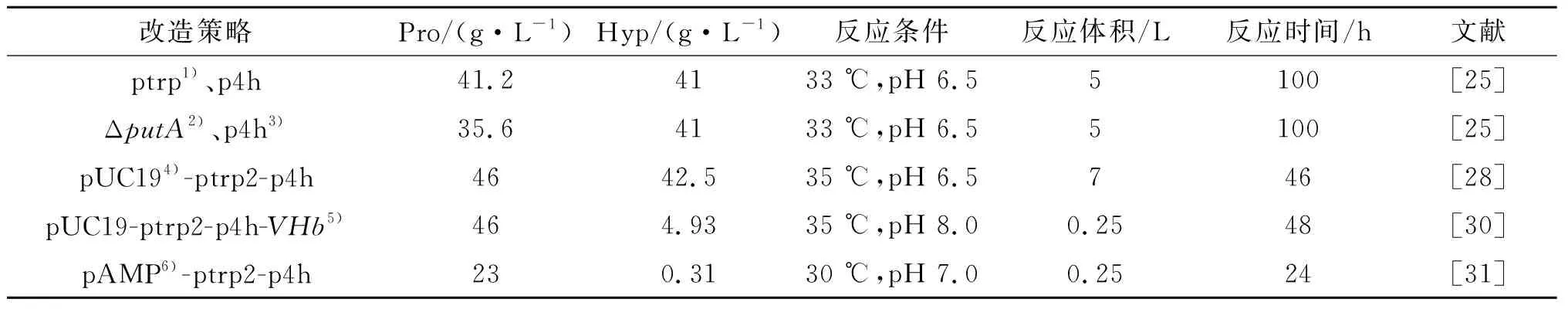
表1 不同全细胞催化法合成羟基脯氨酸的总结Table 1 Summary of hydroxyproline synthetise by different whole cell catalysis
注:1) 色氨酸启动子;2) 敲除L-脯氨酸脱氢酶基因;3) 脯氨酸-4-羟化酶;4) pUC19质粒;5) 透明颤酸血红蛋白基因;6) pAMP质粒.
近年来不少研究者对不同来源的酶做了对比研究.Yi等将不同来源的脯氨酸-4-羟化酶基因克隆至大肠杆菌中,发现来源于指孢囊菌的脯氨酸-4-羟化酶表达的酶活性最高[32].表2是不同来源的脯氨酸4-羟化酶在不同宿主菌中的酶学活性对比,其中酶活为1 min催化脯氨酸生成1 nmol Hyp所需的酶量;比酶活是每毫克湿细胞的酶活.结果显示来源于指孢囊菌的酶在大肠杆菌中表达酶活最高;随后在研究脯氨酸的添加量对产量的影响实验中得出:在添加4 mmol/L的脯氨酸时羟基脯氨酸的质量浓度最高(6.72 g/L).周海岩等为了提高脯氨酸羟基化酶的酶活将来源于根瘤菌(Bradyrhizobiumjaponicum)的反式-4-脯氨酸羟化酶基因p4hBJ(GenBankNO.BAL06808.1)进行序列优化和设计,在大肠杆菌中进行异源表达,全细胞催化反应22 h后,羟基脯氨酸的的质量浓度达到34.86 mg/L[33].
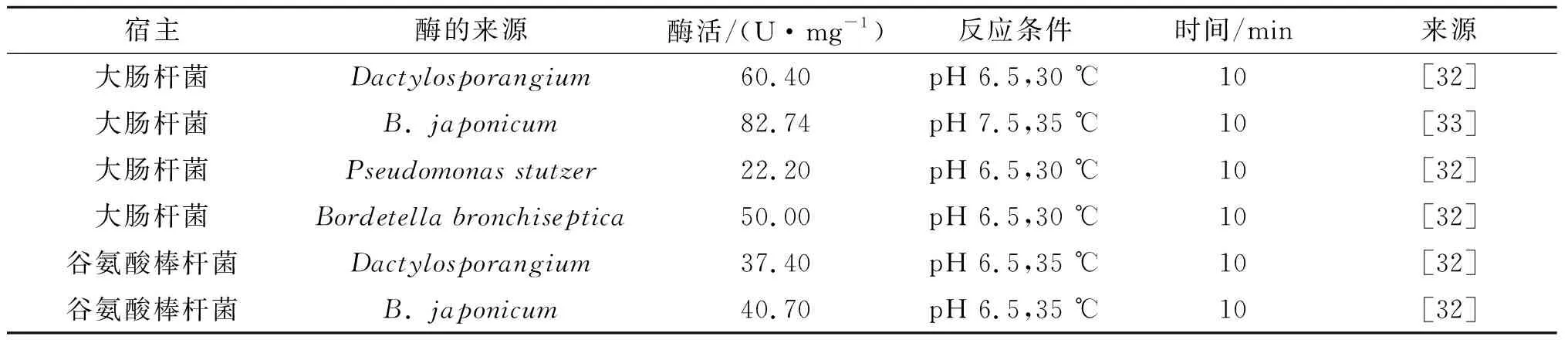
表2 不同来源的脯氨酸-4-羟化酶在不同宿主菌中的活性对比Table 2 Comparison of the activity of proline-4-hydroxylase from different sources in different strains
3.3 直接发酵法
全细胞催化法合成羟基脯氨酸需要额外添加L-脯氨酸做底物,增加了生产成本.在此基础上,研究人员把L-脯氨酸的生物合成途径与反式-4-羟脯氨酸的合成途径结合起来,构建高产羟脯氨酸的工程菌,在不需要额外添加L-脯氨酸的前提下,能够以廉价的葡萄糖为原料,通过液体发酵,在微生物体内进行连续转化生成反式-4-羟脯氨酸,合成途径为

2000年,Shibasaki将脯氨酸-4-羟化酶基因与抗反馈抑制的γ-谷氨酰激酶proB基因和γ-谷氨酰磷酸还原酶proA基因一起在大肠杆菌W1485/ΔputA中表达,以葡萄糖为碳源,发酵96 h产生25 g/L的羟基脯氨酸,由于不用额外添加L-脯氨酸,因此生产成本更低[34].近年来,国内对发酵法生产羟基脯氨酸的研究也日益增多,主要是以大肠杆菌为宿主菌,根据羟基脯氨酸的代谢途径进行改造.2015年胡丹丹等通过对大肠杆菌谷氨酸激酶突变得到突变基因proBA2,降低L-脯氨酸对谷氨酸激酶的反馈抑制作用,通过摇瓶发酵生产反式-4-羟脯氨酸产量为0.22 g/L[35].盛花开等首次将proB,proA,proC和p4h4个基因在同一质粒上表达,在不添加脯氨酸时,摇瓶发酵Hyp的质量浓度为0.16 g/L[36].表3是不同基因工程菌产羟基脯氨酸的含量对比.2017年,王晓姣等以E.coliBL21(DE3)ΔputA为出发菌株,利用基因敲除技术阻断L-谷氨酸到精氨酸的分支途径,增加L-脯氨酸合成代谢流,摇瓶发酵产生312.67 mg/L的羟基脯氨酸,但是在发酵培养基中需要添加脯氨酸和精氨酸,增加了成本[37].
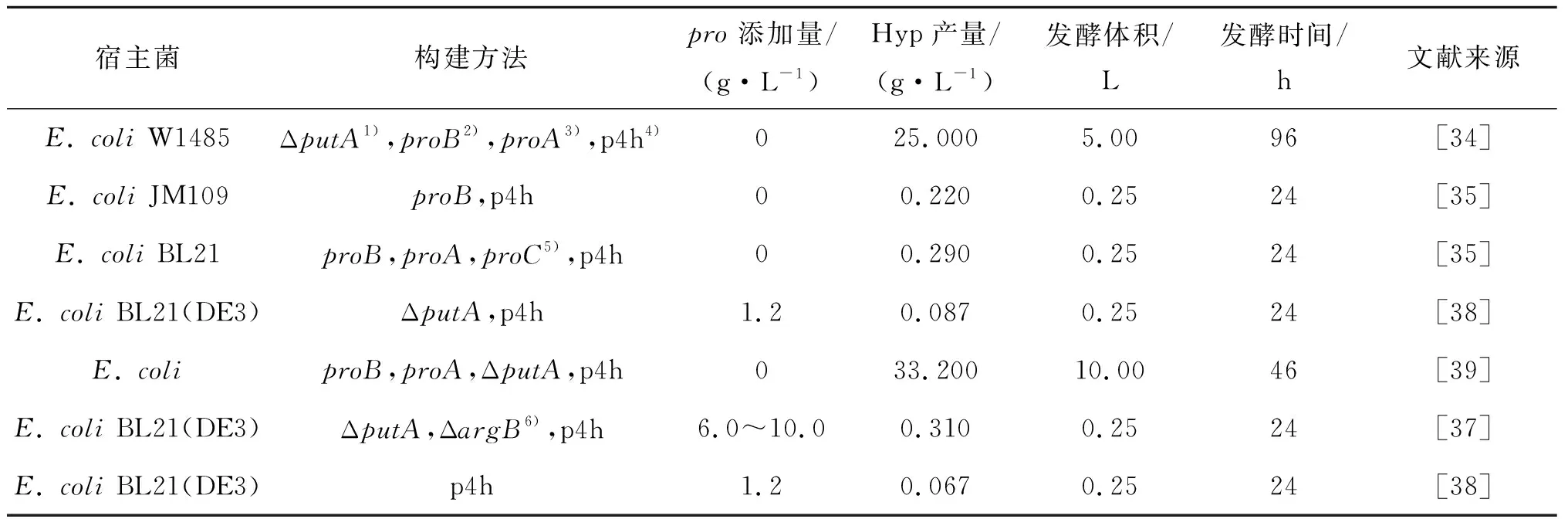
表3 高产羟基脯氨酸的基因工程菌构建方法Table 3 Construction methods of high-yield hydroxyproline genetic engineering bacteria
注:1) L-脯氨酸脱氢酶;2) γ-谷氨酰激酶;3) γ-谷氨酰磷酸还原酶;4) 脯氨酸-4-羟化酶;5) 二氢吡咯羧酸还原酶;6) N-乙酰谷氨酸激酶.
表4总结了羟基脯氨酸三种生产方法.此外,不少研究者以大肠杆菌以外的菌株为羟基脯氨酸的出发菌株进行研究.1995年Serizawa等使用自动筛选系统筛选各种微生物,发现真菌Clonostachyscylindrospora能产生13.8 μg/mL的羟基脯氨酸[40].2014年,研究人员提出将脯氨酸4-羟化酶基因克隆至谷氨酸棒杆菌中生产羟基脯氨酸[32].随后,Falcioni等将来自指孢囊菌的脯氨酸-4-羟化酶基因引入到能产生脯氨酸的谷氨酸棒状杆菌菌株中,利用高密度细胞发酵以葡萄糖为底物,在23 h能生产7.1 g/L的Hyp[41].
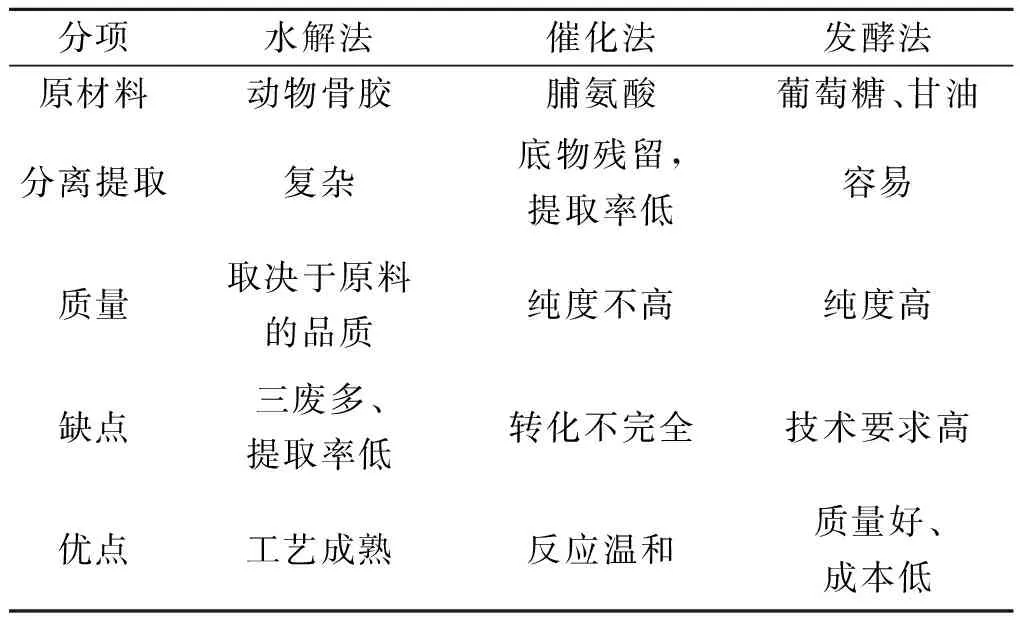
表4 羟基脯氨酸生产方法比较Table 4 Comparison of hydroxyproline production methods
4 结 论
随着羟基脯氨酸应用领域的不断扩展,国内外市场对羟基脯氨酸的需求量逐年增加,对羟基脯氨酸的纯度及其质量要求也相应的提高.目前羟基脯氨酸的生产方法还存在以下问题:1) 脯氨酸羟基化酶催化L-脯氨酸合成羟基脯氨酸时,酶的催化效率低、反应时间长;2) 全细胞催化法需要添加L-脯氨酸,增加生产成本,影响后续的提取分离.因此,可从以下几个方面进行深入研究:1) 完善脯氨酸羟基酶的分子催化机制,以更好地利用蛋白质工程技术提高羟基化酶对底物的特异性和催化活性;2) 利用生物信息学从基因库中筛选具有催化羟基脯氨酸合成的酶,比较其酶性质和合成羟基氨酸的能力,开发催化效率高、稳定性好的酶用于工业生产;3) 利用合成生物学技术控制宿主菌的代谢途径,使更多前体物质流向L-脯氨酸合成途径,从而提高反式-4-羟脯氨酸的产量和转化率.
参考文献:
[1] ANANTHANARAYANAN V S. Structural aspects of hydroxyproline-containing proteins[J].Journal of biomolecular structure & dynamics,1983,1(3):843-855.
[2] GORRES K L, RAINES R T. Prolyl 4-hydroxylase[J].Critical reviews in biochemistry and molecular biology,2010,45(2):106-124.
[3] PHANG J M, LIU W, ZABIRNYK O. Proline metabolism and microenvironmental stress[J].Annual review of nutrition,2010,30(30):441-463.
[4] SCRIVER C R, SMITH R J, PHANG J M. Disorders of proline and hydroxyproline metabolism[J].The metabolic basis of inherited disease,1989,81:577-597.
[5] KIVIRIKKO K I. Urinary excretion of hydroxyproline in health and disease[J].International review of connective tissue research,1970,5:93-163.
[6] RIEDEL T J, JOHNSON L C, KNIGHT J, et al. Structural and biochemical studies of human 4-hydroxy-2-oxoglutarate aldolase: implications for hydroxyproline metabolism in primary hyperoxaluria[J].Plos one,2011,6(10):26021.
[7] ADAMS E, FRANK L. Metabolism of proline and the hydroxyprolines[J].Annual review of biochemistry,1980,49:1005-1061.
[8] KRANE S M. The importance of proline residues in the structure, stability and susceptibility to proteolytic degradation of collagens[J].Amino acids,2008,35(4):703-710.
[9] PRIVALOV P L. Stability of proteins: proteins which do not present a single cooperative system[J].Advances in protein chemistry,1982,35(4):1-104.
[10] BELL E L, EMERLING B M, CHANDEL N S. Mitochondrial regulation of oxygen sensing[J].Mitochondrion,2005,5(5):322-332.
[11] WU G, BAZER F W, BURGHARDT R C, et al. Proline and hydroxyproline metabolism: implications for animal and human nutrition[J].Amino acids,2011,40(4):1053-1063.
[12] NISHINO Y, KOMURASAKI T, YUASA T, et al. Practical large-scale synthesis of the 2-aminomethylpyrrolidin-4-ylthio-containing side chain of the novel carbapenem antibiotic doripenem[J].Organic process research & development,2003,7(5):649-654.
[13] 张义凤,陈昊,彭久合,等.碳青霉烯类抗生素厄他培南的合成[J].中国药科大学学报,2007,38(4):305-310.
[14] 郑子新,靳继德,徐琪寿.富含谷氨酰胺和支链氨基酸的肠外制剂对创伤大鼠的效用[J].感染、炎症、修复,2002,3(1):40-42.
[15] ALCAIDE B, ALMENDROS P. The direct catalytic asymmetric aldol reaction[J].European journal of organic chemistry,2002(10):1595-1601.
[16] 陈玲,李扬,农蕊瑜,等.反式-4-羟基-L-脯氨酸手性固定相的制备及性能评价[J].分析试验室,2015,34(12):1365-1369.
[17] PHANG J M, DONALD S P, PANDHARE J, et al. The metabolism of proline, a stress substrate, modulates carcinogenic pathways[J].Amino acids,2008,35(4):681-690.
[18] TAKEKOSHI Y, TAKAHASHI T, OONUMA T. Skin care product containing hydroxyproline or N-acylated hydroxyproline derivatives: EP 1166767 A3[P].2004-01-28.
[19] EL-SISI A E, ELFERT A A, EL-SAYAD M, et al. A randomized controlled study of the effect of at1 antagonist on fibrosis markers in hcv egyptian patients[J].Journal of the acoustical society of America,2012,37(1):174-174.
[20] SILVA K L G L, COELHO R A P, MARINS J C B, et al. Efeitos do Alongamento sobre os níveis de hidroxiprolina em atiradores do tiro de guerra[J].Fitness & performance journal,2005,4(6):348-351.
[21] NOGUEIRA A D C, PASSOS C T, VALE R G D S, et al. Sobrecarga cardiovascular nos tipos de ação muscular dos exercícios resistidos[J].Fitness & performance journal,2007,6(2):105-110.
[22] 李娟,陈舜胜.从明胶水解液中分离制备L-羟脯氨酸和L-脯氨酸[J].现代食品科技,2006,22(4):151-152.
[23] BALDWIN J E, FIELD R A, LAWRENCE C C, et al. Substrate specificity of proline-4-hydroxylase: chemical and enzymatic synthesis of 2S, 3R, 4S-epoxyproline[J].Tetrahedron letters,1994,35(26):4649-4652.
[24] SHIBASAKI T, MORI H, CHIBA S, et al. Microbial proline 4-hydroxylase screening and gene cloning[J].Applied and environmental microbiology,1999,65(9):4028-4031.
[25] SHIBASAKI T, MORI H, OZAKI A. Enzymatic production oftrans-4-hydroxy-L-proline by regio- and stereospecific hydroxylation of L-proline[J].Bioscience biotechnology & biochemistry,2000,64(4):746-750.
[26] 张琳琳,张庆,李莉,等.反式-4-羟基-L-脯氨酸羟化酶基因密码子优化及表达[J].河北师范大学学报(自然科学版),2017,41(4):341-347.
[27] 刘笑尘,高爽,许伟,等.产反式-4-羟脯氨酸重组大肠杆菌的构建及优化[J].江苏农业科学,2017,45(13):19-23.
[28] 刘合栋,袁春伟,张震宇.高产反式-4-羟脯氨酸重组大肠杆菌的构建以及发酵条件优化[J/OL].[2013-01-23].中国科技论文在线,http://www.paper.edu.cn/releasepaper/content/201301-989.
[29] 袁春伟,何艳春,张胜利,等.重组大肠杆菌BL21(pUC19 Hyp)产羟脯氨酸的补料分批培养[J].生物加工过程,2014,12(4):43-48.
[30] 张震宇,刘合栋,张胜利.一种使含有重组DNA的反式-4-羟脯氨酸生物合成系统活性增强的方法:CN 104278047 A[P].2013-07-04.
[31] 张震宇,刘合栋,袁春伟.一种利用重组大肠杆菌发酵生产L-4-羟脯氨酸的方法:CN 103509813 A[P].2012-06-20.
[32] YI Y, SHENG H, LI Z, et al. Biosynthesis oftrans-4-hydroxyproline by recombinant strains ofCorynebacteriumglutamicumandEscherichiacoli[J].BMC biotechnology,2014,14(1):44.
[33] 周海岩,李会帅,王培,等.L-脯氨酸-4-羟化酶的异源表达和合成反式-4-羟基-L-脯氨酸的条件优化[J].工业微生物,2017,47(1):1-9.
[34] SHIBASAKI T, HASHIMOTO S, MORI H, et al. Construction of a novel hydroxyproline-producing recombinantEscherichiacoliby introducing a proline 4-hydroxylase gene[J].Journal of bioscience & bioengineering,2000,90(5):522-525.
[35] 胡丹丹,王付才,张震宇,等.无外源脯氨酸发酵产反式-4-羟脯氨酸重组大肠杆菌的构建及发酵优化[J].食品工业科技,2015,36(20):168-174.
[36] 盛花开,衣玉兰,李志敏,等.重组大肠杆菌多基因串联表达合成反式-4-羟基-L-脯氨酸[J].生物技术,2016,26(1):81-86.
[37] 王晓姣,张震宇,孙付保,等.精氨酸缺陷型菌株发酵生产反式-4-羟脯氨酸[J].食品与发酵工业,2017,43(1):24-30.
[38] 张胜利,林凡,刘合栋,等.putA与vgb基因对反式-4-羟基-L-脯氨酸产量的影响[J].食品与生物技术学报,2016,35(12):1307-1316.
[39] 储消和,吴黎诚.一种生产反式-4-羟基-L-脯氨酸的发酵工艺:CN 201510899082.2[P].2015-12-09.
[40] SERIZAWA N, MATSUOKA T, HOSOYA T, et al. Fermentative production oftrans-4-hydroxy-L-proline byClonostachyscylindrospora[J].Journal of the agricultural chemical society of Japan,1995,59(3):555-557.
[41] FALCIONI F, BüHLER B, SCHMID A. Efficient hydroxyproline production from glucose in minimal media byCorynebacteriumglutamicum[J].Biotechnology & bioengineering,2015,112(2):322-330.
猜你喜欢
中华实用诊断与治疗杂志(2022年2期)2022-09-02
纺织标准与质量(2022年1期)2022-07-12
中国生殖健康(2020年5期)2021-01-18
热带作物学报(2020年9期)2020-10-29
中国现代医生(2020年12期)2020-07-04
中国生殖健康(2018年5期)2018-11-06
中国酿造(2018年7期)2018-08-10
食品与机械(2018年5期)2018-07-16
中国民族医药杂志(2016年5期)2016-05-09
食品安全导刊(2016年15期)2016-03-27
