
大鼠脊髓损伤后脊髓组织中上调差异基因的生物信息学分析
2019-06-28 02:16钟占琼薛清彩易晓红黄聪于海艳刘伟伟陈继兰夏军张晓
中国康复理论与实践 2019年6期
钟占琼,薛清彩,易晓红,黄聪,于海艳,刘伟伟,陈继兰,夏军,张晓
1.成都中医药大学基础医学院,四川成都市611137;2.成都医学院基础医学院,四川成都市610500
作为中枢神经系统的脊髓组织在机体的生命活动中发挥重要作用,脊髓的剧烈损伤伴随着严重的上行和下行通路的传导障碍。随着交通的发达和极限运动的流行,脊髓损伤(spinal cord injury,SCI)的发生率持续增加。据最新统计,脊髓损伤影响全球人数超过2704万[1]。物理性损伤导致损伤段脊髓出血、肿胀等,神经元和神经胶质细胞缺血、缺氧、坏死。随着脊髓水肿和炎症的发生,神经元和神经胶质细胞的坏死、凋亡向损伤中心以外的组织扩散,神经胶质瘢痕的形成可以稳定继发性损伤的扩散,但也阻止了轴突的再生[2]。炎症反应、水肿、缺血、缺氧、兴奋性毒性、自由基损伤、脂质过氧化、凋亡、神经胶质瘢痕的形成等[3]继发性损伤严重影响神经可塑性和功能恢复。
目前,有大量实验研究显示脊髓损伤后出现基因上调或下降,如白细胞介素-10(interleukin-10,IL-10)、人原肌球蛋白α4(tropomyosin 4,TPM4)、B淋巴细胞瘤-2相关蛋白X(B-cell lymphoma-2 associated X protein,Bax)、B淋巴细胞瘤-2(B-cell lymphoma-2,Bcl-2)、血管内皮生长因子(vascular endothelial growth factor,VEGF)、半胱氨酸蛋白酶-3(Caspase-3,Casp-3)、水通道蛋白4(aquaporin 4,AQP4)[4-5]等。然而,大多研究主要从单个基因或蛋白与脊髓损伤后细胞的炎症、凋亡和水肿等多种改变相关联进行研究,缺少系统分析。
近年来,借助生物技术手段从整体上探索基因组在疾病中的作用及规律越来越受到重视。基因芯片技术正是基于这种手段通过杂交测序方法,将已知序列的脱氧核糖核酸(deoxyribonucleic acid,DNA)探针与待测的经标记的DNA样品杂交,同时检测上万种基因。具有高通量和全面特性的基因芯片技术,现已经广泛用于DNA测序、基因表达分析与基因调控机理的研究。目前,基因分析运用于神经系统疾病的研究越来越多[6],例如阿尔茨海默病[7]、孤独症[8]和帕金森综合征[9]。此外,基因分析也运用于脑损伤和脊髓损伤等疾病[10]。
本课题前期构建大鼠脊髓损伤模型,取脊髓损伤段组织用基因芯片的技术检测基因表达。结合课题组前期结果,本文主要分析在检测出差异表达基因中的上调基因部分,采用基因本体论(gene ontology,GO)和基因组百科全书数据库(Kyoto Encyclopedia of Genes and Genomes,KEGG)分析差异上调基因的功能定位与通路,主要从生物学途径(biological process,BP)、细胞组件(cellular component,CC)和分子功能(molecular function,MF)三方面进行分析。
1 材料与方法
1.1 实验动物与分组
健康成年雌性Sprague-Dawley大鼠6只(成都达硕公司),体质量(200±20)g,单笼饲养,环境温度20~28℃。实验动物随机分为对照组(sham组)和脊髓损伤组(SCI组),正常光照和饮食。
1.2 动物模型制备
3.6 %水合氯醛10 ml/kg腹腔注射麻醉实验大鼠。俯卧位固定、消毒。定位T10棘突,常规行椎板切除术,暴露脊髓。将10 g金属棒从30 mm处自由下落,造成脊髓钝伤模型(脊髓损伤组)。对照组只剥离椎板,暴露脊髓但不损伤脊髓。术后逐层缝合伤口,每天给予大鼠肌肉注射青霉素80 000 IU/kg,持续3 d。大鼠正常进食、饮水,独笼饲养。每天两次协助排尿,直至大鼠恢复自主排尿。
在术后第3天,以损伤段脊髓为中心点,取脊髓组织1 cm,-80℃冰箱保存。
1.3 基因芯片的检测
取脊髓组织提取RNA后,进行基因芯片检测分析,该部分交于上海其明公司进行操作。采用昂飞基因芯片2.0版本(Affymetrix GeneChip Gene 2.0芯片)进行检测分析。基因芯片用操作软件(GeneChip Operating Software,GCOS)扫描得到的图像,采用Agilent GeneSpring GX software(version 11.5.1)分析,所得的表达数据均已进行标准化处理。用差异基因倍数改变的筛选方法,以检测到的样本信号强度比值上调或下调大于1.5倍界定为差异表达基因。采用MultiExperiment Viewer(version 4.9.0)进行分级群聚的聚类分析,并对差异基因中的上调基因部分深入进行GO(http://www.geneontology.org/)分析及KEGG(http:www.genome.jp/kegg/)信号通路分析[11]。
2 结果
2.1 样本质量
将提取脊髓组织总RNA上样于1%琼脂糖凝胶孔中,120 V电压5 min。利用紫外分光光度仪测量RNA浓度,并通过A260/A280值初步判断RNA质量报告(表1)。釆用RNA变性琼脂糖凝胶电泳法检测RNA完整性,可见明显的28S和18S条带,未见其他明显杂带,无污染,未降解,可用于下一步分析(图1)。
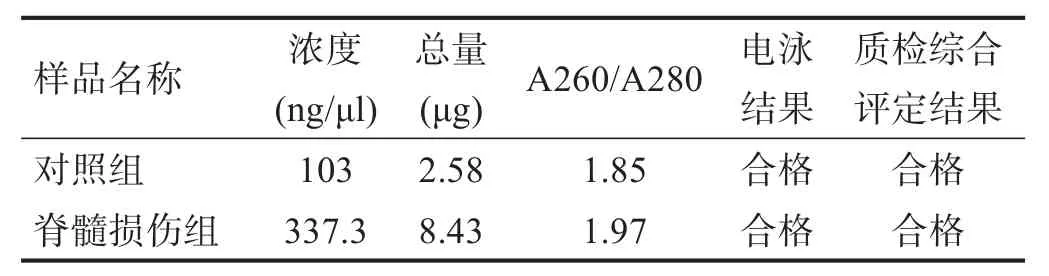
表1 样本总RNA检测结果

图1 脊髓组织琼脂糖电泳结果
2.2 差异表达基因
两组差异表达基因4222个,其中上调基因1874个,下调基因2348个。利用Cluster软件导入数据,制作热图(图2)。

图2 脊髓损伤组与对照组差异基因热图
2.3 生物信息学分析
2.3.1 生物学途径
根据差异基因数量排名前15位的生物学途径分别是:参与负调控凋亡过程(negative regulation of apoptotic process)478个,对药物的反应(response to drug)462个,蛋白质磷酸化(protein phosphorylation)353个,负调控细胞增殖(negative regulation of cell proliferation)308个,细胞黏附(cell adhesion)260个,正调控凋亡过程(positive regulation of apoptotic process)258个,对缺氧的反应(response to hypoxia)242个,细胞内信号转导(intracellular signal transduction)236个,对有机环状化合物的反应(response to organic cyclic compound)230个,代谢过程(metabolic process)230个,免疫反应(immune response)215个,对脂多糖(response to lipopolysaccharide)的反应211个,心脏发育(heart development)191个,炎症反应(inflammatory response)189个,老化(aging)189个(图3A)。
根据富集倍数排名前15位的生物学途径分别是:模式识别受体信号通路(pattern recognition receptor signaling pathway)、吞噬溶酶体组装(phagolysosome assembly)、细胞-基底连接组件(cell-substrate junction assembly)、参与免疫反应的嗜中性粒细胞活化(neutrophil activation involved in immune response)、参与免疫反应的白细胞活化(leukocyte activation involved in immune response)、神经节苷脂分解代谢过程(ganglioside catabolic process)、寡糖分解代谢过程(oligosaccharide catabolic process)、铜离子的解毒(detoxification of copper ion)、抗原加工和通过主要组织相容性复合体I类呈递内源性肽抗原(antigen processing and presentation of endogenous peptide antigen via major histocompatibility complex class I)、检测脂多糖(detection of lipopolysaccharide)、Toll样受体7信号通路(Toll-like receptor 7 signaling pathway)、Fc-γ受体信号传导途径(Fc-gamma receptor signaling pathway)、信号传导及转录激活因子蛋白酪氨酸磷酸化的正调控(positive regulation of tyrosine phosphorylation of signal transducer and activator of transcription protein)、抗原加工和通过主要组织相容性复合体I类呈递外源性肽抗原(antigen processing and presentation of exogenous peptide antigen via major histocompatibility complex class I)、负向调节病毒进入宿主细胞(negative regulation of viral entry into host cell)(图 3B)。
根据富集积分(enrichment score)排名前15位的生物学途径分别是:先天免疫反应(innate immune response)、对脂多糖的反应(response to lipopolysaccharide)、炎症反应(inflammatory response)、对有机环状化合物的反应(response to organic cyclic compound)、对药物的反应(response to drug)、正调控凋亡过程(positive regulation of apoptotic process)、细胞黏附(cell adhesion)、负调控凋亡过程(negative regulation of apoptotic process)、细胞迁移的正调控(positive regulation of cell migration)、细胞对脂多糖的反应(cellular response to lipopolysaccharide)、整合素介导的信号传导途径(integrin-mediated signaling pathway)、正调节IκB 激酶/核因子-κB级联反应(positive regulation of I-kappaB kinase/nuclear factor-kappaB cascade)、老化(aging)、蛋白质磷酸化(protein phosphorylation)、免疫反应(immune response)(图 3C)。
本研究在查阅大量相关文献的基础上,根据试验的目的和计划,着重探讨大花序桉茎段外植体消毒及其芽诱导组织培养技术,采用正交设计或完全组合设计安排试验,探讨外源激素的添加效果,筛选最有利的内外源激素的组合和比例,筛选出最优技术参数组合,从外源激素的协调上建立大花序桉最优组培快繁体系。
2.3.2 细胞组分
根据差异基因数量排名前15位的细胞组件分别是:细胞质(cytoplasm)455个、细胞核(nucleus)432个、细胞外囊泡外泌体(extracellular vesicular exosome)405个、微膜(integral to membrane)319个、质膜(plasma membrane)300个、细胞膜(membrane)271个、细胞外空间(extracellular space)173个、核仁(nucleolus)158个、细胞成分(cellular_component)150个、细胞质溶胶(cytosol)142个、细胞内(intracellular)114个、细胞表面(cell surface)108个、线粒体(mitochondrion)93个、局部性黏附(focal adhesion)93个、内质网(endoplasmic reticulum)84个 (图4A)。

图3 上调基因的分子生物学途径分析图
根据富集度排名前15位的细胞组分分别是:肌动蛋白相关蛋白2/3复合体(actin-related protein-2/3 complex,Arp2/3 protein complex)、炎性小体3复合体(NOD-like receptor pyrin domain containing 3 inflamma-some complex,NLRP3 complex)、凝聚核染色体外着丝粒(condensed nuclear chromosome outer kinetochore)、异常糖链糖蛋白复合体(transporter of antigenic peptides,TAP complex)、转化生长因子β受体同型二聚复合物(transforming growth factor beta receptor homodimeric complex,TGF-βr homoderic complex)、炎性小体1复合体(NOD-like receptor pyrin domain containing 1 inflammasome complex,NLRP1 complex)、胞质分裂相关蛋白复合体(centralspindlin complex)、I型胶原蛋白(collagen type I)、早期内体腔(early endosome lumen)、B细胞淋巴瘤3基因/核转录因子2复合物(B-cell lymphoma 3/nuclear factor-kappaB 2 complex,Bcl3/NF-κB2 complex)、整联蛋白α1β1复合物(alpha1-beta1 integrin complex,α1β1 integrin complex)、整联蛋白αLβ2复 合 物 (alphaL-beta2 integrin complex,αLβ2 integrin complex)、Toll样受体2-Toll样受体6蛋白复合物(Tolllike receptor 2-Toll-like receptor 6 protein complex)、β-连环蛋白-转录因子7类似物2复合物(beta-catenintranscription factor 7-like-2 complex,β-catenin-TCF7L2 complex)、α9-β1-整合素-去整合素和金属蛋白酶域-8复合物(alpha9-beta1 integrin-a disintegrin and metalloprotease complex, α9-β1 intergin-ADAM8 complex)(图 4B)。
根据富集积分排名前15位的细胞组件分别是:细胞外囊泡外泌体(extracellular vesicular exosome)、细胞质(cytoplasm)、细胞核(nucleus)、细胞膜(membrane)、细胞外空间(extracellular space)、质膜(plasma membrane)、细胞表面(cell surface)、局部性黏附(focal adhesion)、积分膜(integral to membrane)、细胞外基质(extracellular matrix)、细胞质溶胶(cytosol)、核仁(nucleolus)、细胞内(intracellular)、溶酶体(lysosome)、细胞成分(cellular component)(图4C)。
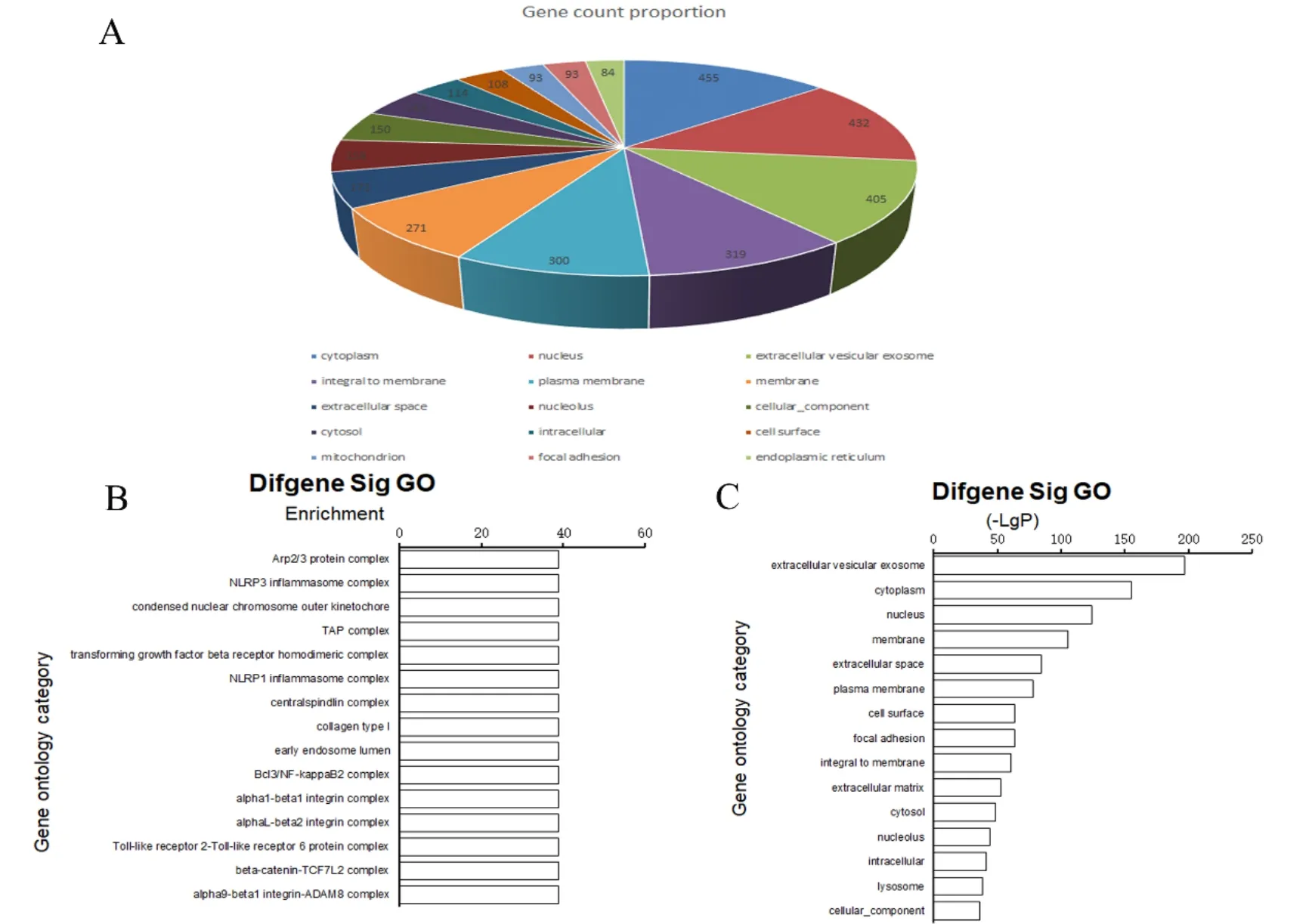
图4 上调基因的细胞组分分析图
2.3.3 分子功能
按照差异基因数量排名前15位的分子功能分别是:与分子功能相关(molecular function)189个,与蛋白质结合(protein binding)154个,与腺嘌呤核苷三磷酸结合(Adenosine triphosphate binding,ATP binding)150个,与DNA结合(DNA binding)96个,与金属离子结合(metal ion binding)93个,与锌离子结合(zinc ion binding)93个,与蛋白质同源二聚化活性(protein homodimerization activity)79个,与相同的蛋白结合(identical protein binding)72个,与钙离子结合(calci-um ion binding)70个,与蛋白质异源二聚化活性(protein heterodimerization activity)69个,与蛋白激酶结合(protein kinase binding)60个,与序列特异性DNA结合转录因子活性(sequence-specific DNA binding transcription factor activity)56个,与蛋白复合物结合(protein complex binding)50个,与蛋白质丝氨酸/苏氨酸激酶活性(protein serine/threonine kinase activity)42个,与碳水化合物结合(carbohydrate binding)42个(图 5A)。
根据富集度排名前15位的分子功能分别是:趋化因子受体3结合(C-X-C chemokine receptor type 3,CXCR3 chemokine receptor binding)、脂多糖受体活性(lipopolysaccharide receptor activity)、脂磷壁酸结合(lipoteichoic acid binding)、β-N-乙酰己糖胺酶活性(beta-N-acetylhexosaminidase activity)、前胶原-赖氨酸 5-双加氧酶活性(procollagen-lysine 5-dioxygenase activity)、嘌呤核苷酸结合(purine nucleotide binding)、IgG受体活性(IgG receptor activity)、嘌呤核苷碱基结合(purine nucleobase binding)、磷酸二酯酶I活性(phosphodiesterase I activity)、芳基烃受体活性(aryl hydrocarbon receptor activity)、白细胞介素-10受体活性(interleukin-10 receptor activity)、花生四烯酸11,12-环氧化酶活性(arachidonic acid 11,12-epoxygenase activity)、肽抗原转运ATP酶活性(peptide antigen-transporting ATPase activity)、凋亡蛋白酶激活剂活性(apoptotic protease activator activity)、主要组织相容性复合体Ib类蛋白结合(major histocompatibility complex class Ib protein binding)(图 5B)。
根据富集积分排名前15位的分子功能分别是:分子功能(molecular function)、蛋白质结合(protein binding)、ATP结合、蛋白质异二聚化活性(protein heterodimerization activity)、蛋白激酶结合(protein kinase binding)、相同的蛋白结合(identical protein binding)、蛋白质同源二聚化活性(protein homodimerization activity)、DNA结合、整合素结合(integrin binding)、碳水化合物结合(carbohydrate binding)、肝素结合(heparin binding)、蛋白复合物结合(protein complex binding)、锌离子结合(zinc ion binding)、钙离子结合(calcium ion binding)、金属离子结合(metal ion binding)(图 5C)。
图5 上调基因的分子功能分析图
2.4 信号通路
脊髓损伤组上调显著的信号通路共103个,根据P值的高低,前15种信号通路分别是:破骨细胞分化(osteoclast differentiation)、吞噬体(phagosome)、人类嗜T细胞病毒I型感染(human T cell leukemia virus type-I,HTLV-I infection)、溶酶体(lysosome)、Toll样受体信号通路(Toll-like receptor signaling pathway)、细胞因子-细胞因子受体相互作用(cytokine-cytokine receptor interaction)、磷脂酰肌醇三激酶-蛋白激酶B信号通路(phosphatidylinositol 3-kinase protein kinase B signaling pathway,PI3K-Akt signaling pathway)、系统性红斑狼疮(systemic lupus erythematosus)、趋化因子信号通路(chemokine signaling pathway)、局部黏附(focal adhesion)、结核(tuberculosis)、利什曼病(leishmaniasis)、甲型流感(influenza A)、细胞外基质-受体相互作用(extracellular matrix receptor interaction)、弓形体病(toxoplasmosis)(图 6)。

图6 上调基因的信号通路
3 讨论
脊髓损伤后,原发性损伤和继发性损伤导致感觉和运动功能障碍,严重影响家庭的生活质量,因此,深入研究治疗脊髓损伤具有重大的意义。当前研究的主要目标是找出影响脊髓损伤后功能恢复的关键基因和蛋白。本研究系统检测了脊髓损伤后基因的表达变化,采用生物信息学的方法分析差异上调基因的主要功能、定位和通路,为后续找准关键靶点指明方向。
当前,众多脊髓损伤实验研究根据不同的实验目的和要求,损伤实验动物T10脊髓段,并选取对应损伤点1 cm脊髓组织用于研究[12],或损伤T9~T10胸椎,并选取对应脊髓段(T8~T10)用于后续的基因水平表达或蛋白的研究[13]。本课题组前期预实验结果显示,脊髓损伤后同一基因在脊髓损伤段和损伤上段、损伤下段表达有差异,为了得到更为准确的结果,本研究构建脊髓钝挫伤模型,根据动物脊髓对应的节段范围,选取以损伤段为中心的损伤段脊髓(约1 cm)用以研究。
本研究采用Affymetrix GeneChip Gene 2.0芯片检测脊髓损伤组和对照组在损伤后脊髓损伤段组织的差异表达基因,结果显示上调或下调1.5倍的差异基因有4222个,包括1874个上调基因和2348个下调基因。脊髓损伤后有大量基因发生差异性变化,尤其是下调基因占多数。以往的实验研究结果显示,与神经痛相关的环指蛋白34(ring finger protein 34,RNF34)和垂体腺苷酸环化酶激活肽(pituitary adenylate cyclase activating polypeptide,PACAP)基因水平在脊髓损伤后增加[14],与炎症相关的白细胞介素-6、白血病抑制因子、白细胞介素-11等基因水平在脊髓损伤后增加[15]。与凋亡有关的锌指蛋白38基因水平下调[16],而哺乳动物无菌-20样激酶1(mammalian ste20-like kinase 1,Mst1)在野生型小鼠损伤的脊髓组织中表达升高[17];在脊髓完全损伤的模型中,脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)、胶质细胞源性神经营养因子(glial cell-derived neurotrophic factor,GDNF)的表达降低[18]。在脊髓损伤后48 h,AQP4表达降低[19]。结合本实验芯片结果,我们认为脊髓损伤后有差异基因上调或者下调,这与基因参与脊髓损伤后功能相一致。此外,同一个基因在不同的损伤阶段和不同的组织中可能会出现不同的结果,甚至相反的结果。实验研究表明,AQP4在脊髓损伤后的急性期和慢性期表达不一样,与损伤后不同时间段水肿有关[20]。
根据芯片的检测结果进行初步分析后,将1874个上调基因按照生物学途径进行分类,分别从生物学过程分类排序、富集倍数和富集分数选取排名前15位的生物学途径。综合三项结果分析,脊髓损伤后上调基因大多参与负调控凋亡过程、蛋白质磷酸化、负调控细胞增殖、细胞迁移、细胞黏附、正调控凋亡过程、对缺氧的反应、细胞内信号转导、有机环状化合物的反应、代谢过程、免疫反应、抗原加工和通过主要组织相容性复合体I类呈递内源性肽抗原、对脂多糖的反应、心脏发育、炎症反应、老化、对药物的反应、神经节苷脂分解代谢过程,提示脊髓损伤过程中出现大量涉及细胞基本代谢、凋亡、炎症和免疫反应的基因表达上调,可能与脊髓损伤后神经元和神经胶质细胞变性、死亡、凋亡等密切相关,并引发炎症和免疫应答反应,因此影响细胞功能的过程,是一个多基因参与的复杂过程。脊髓损伤后炎症反应引发机体的功能障碍,予芬维A胺、沉默信息调节因子2相关酶1(silent information regulator 2,SIRT1)等都有助于减轻炎症,促进运动功能的恢复[21]。
从细胞组分定位结果分析,脊髓损伤后脊髓组织中1874个上调基因,大部分定位在细胞质、细胞核、细胞外囊泡外泌体、微膜、质膜、细胞膜、细胞外、核仁、细胞成分、细胞内、线粒体。根据富集度和富积分数选取排名靠前的细胞组分,主要是肌动蛋白相关蛋白2/3复合体、炎性小体3复合体、异常糖链糖蛋白复合体、转化生长因子β受体同型二聚复合物、炎性小体1复合体、胞质分裂相关蛋白复合体、B细胞淋巴瘤3基因/核转录因子2复合物、整联蛋白αLβ2复合物、Toll样受体2-Toll样受体6蛋白复合物、β-连环蛋白-转录因子7类似物2复合物、α9-β1-整合素-去整合素和金属蛋白酶域-8复合物等。说明脊髓损伤后亚急性期,定位于以上亚细胞器的基因较为敏感甚至直接受到损害,诱导免疫应答、炎性反应、缺血缺氧等。提示在脊髓损伤的不同阶段实施不同治疗措施,或许对脊髓损伤的急性期和慢性期有一定的作用,但仍需要进一步研究证实。
从差异基因数量、富集倍数和富集分数研究上调基因的分子功能,结果显示,排名靠前的基因功能主要与细胞成分、离子以及生物活性因子等结合,如分子功能、ATP结合、嘌呤核苷酸结合、DNA结合、嘌呤核苷碱基结合、蛋白复合物结合、金属离子结合、锌离子结合、钙离子结合、蛋白激酶结合、整合素结合、碳水化合物结合、肝素结合、脂磷壁酸等;与多种细胞酶或受体活性等有关,如β-N-乙酰己糖胺酶活性、前胶原-赖氨酸5-双加氧酶活性、磷酸二酯酶I活性、花生四烯酸11,12-环氧化酶活性、肽抗原转运ATP酶活性、蛋白激酶活性、凋亡蛋白酶激活剂活性、趋化因子受体3、脂多糖受体活性、IgG受体活性、芳基烃受体活性、白细胞介素-10受体活性等。提示在脊髓损伤后,脊髓组织中上调的功能基因与脊髓中神经元和神经胶质细胞的功能受损或缺失有关系,并发现有与凋亡、趋化因子和炎症等相关的受体相关的上调基因,进一步说明脊髓损伤后在急性期和亚急性期有炎性因子的浸润,继发性引发细胞功能障碍。我们推测这些基因表达差异可能与脊髓运动功能和感觉功能缺损密切相关,但还有待进一步的证实。
将上调的差异基因进行信号通路分析,参与的通路共有103个,但主要有破骨细胞分化、吞噬、溶酶体、Toll样受体信号通路、细胞因子-细胞因子受体相互作用、PI3K-Akt信号通路、趋化因子信号通路,结果提示,在脊髓损伤后有多条信号通路发生显著改变,涉及到多种细胞内生物学过程,找出信号通路中的关键因子对脊髓损伤后的治疗有重要意义。有实验研究表明,纤维母细胞生长因子10(fibroblast growth factor 10,FGF10)可以通过减少细胞凋亡以及通过PI3K/Akt途径修复神经突,改善脊髓损伤后的功能[22]。
综上所述,本研究通过采用基因芯片技术,研究脊髓损伤组和对照组脊髓组织中的差异表达基因,进一步从生物学途径、细胞组分和分子功能三个方面分析上调差异表达基因,结果提示,在脊髓损伤急性期上调差异基因变化参与多条信号通路,与细胞凋亡、炎性、免疫应答等有关。机体发生脊髓损伤后,发生一系列多基因协同参与调控的复杂过程,对差异表达的基因进行系统分析有助于阐明脊髓损伤后发生变化的生物学机制,为后续研究脊髓损伤提供理论依据。
猜你喜欢
军事文摘(2022年8期)2022-11-03
农业工程学报(2022年13期)2022-10-09
九江学院学报(自然科学版)(2022年2期)2022-07-02
现代仪器与医疗(2022年1期)2022-04-19
作物学报(2022年3期)2022-01-22
中华养生保健(2020年5期)2020-11-16
中华养生保健(2020年5期)2020-11-16
西华大学学报(自然科学版)(2020年6期)2020-10-15
中华肩肘外科电子杂志(2019年4期)2019-08-24
电子制作(2019年7期)2019-04-25
