
2种烷基多环芳烃对仿刺参CYP450和p53基因表达的影响研究
2019-06-10 02:11李香魏海峰1刘长发宋雪赵肖依赵雨朦夏宁霍玉洁
生态毒理学报 2019年1期
李香,魏海峰1,,*,刘长发,宋雪,赵肖依,赵雨朦,夏宁,霍玉洁
1. 大连海洋大学,农业农村部北方海水增养殖重点实验室,大连 116023 2. 大连海洋大学,海洋科技与环境学院,大连 116023 3. 大连海洋大学,近岸海洋环境科学与技术辽宁省高校重点实验室,大连 116023
近年来由于突发海洋溢油事故和沿海石油化工业的发展,大量石油烃类污染物通过各种途径进入海洋中,成为潜在的危害我国海洋生态环境安全的重要因素之一。石油毒性主要来源于其中的多环芳烃(PAHs)组分,多环芳烃类约占石油组分的1%~6%,其中80%~90%为烷基化多环芳烃[1]。前期国内外大量研究多关注16种优先控制的多环芳烃,而对多环芳烃的衍生物关注极少,这可能导致由多环芳烃衍生物造成的生态风险被低估[2]。菲和蒽是最具代表性的三环多环芳烃,在石油中含量较高,关于其毒性的研究报道较多[3-4]。3-甲基菲和2-甲基蒽是菲和蒽的烷基化衍生物,国内外关于这2种衍生物毒性的报道较少,有必要对其毒性效应进行研究。
细胞色素P450(Cytochrome P450, CYP450)和p53基因都是常用的分子生物标志物,它们均在外源物质的代谢和解毒方面起重要作用[5-6]。CYP450是生物体内含有多种超家族CYP450血红蛋白或相同结构域的酶系,对许多外源及内源化合物在生物体内的生物转化具有重要的作用[7]。环境中的有机污染物进入生物体内后,可诱导或抑制细胞色素P450酶系CYP活力显著增加或降低,利用生物代谢过程中CYP含量或活力与污染物毒性之间的响应关系,可将CYP450作为毒物毒性的生物标记物,进行环境污染的早期诊断。但由于大多数P450酶的含量很低且不稳定,直接分离纯化P450酶非常困难,因此在克隆P450基因的基础上,研究多环芳烃类污染物对CYP450基因表达的影响[8-9]。抑癌基因p53在细胞凋亡中起着重要作用,DNA损伤以及不正常的细胞增殖信号都会引起P53蛋白的激活[10],由于其功能的多样性及与肿瘤发生的高度相关性,研究环境有毒化合物对p53基因的损失对阐明化合物遗传毒作用、分子机制以及风险评价具有重要的科学意义[6]。
仿刺参(Apostichopusjaponicus)是黄、渤海重要底栖生物类群,主要摄食底泥中细菌、底栖硅藻和有机质碎屑[11]。作为我国重要海水增养殖品种之一,其生物学数据基本完备、并且取材方便,是海洋环境污染物生物毒性评价的理想受试生物[12]。林芳等[13]和靳非等[2]分别进行了多环芳烃类污染物对翡翠贻贝(Pernaviridis)胚胎和河鲀(Takifugurubripes)幼鱼的毒理学研究。相比贝类、鱼类而言,国内外对于多环芳烃类污染物对海参的毒理学效应研究较少。因此,本文选择仿刺参(A.japonicus)为受试生物,设置3种不同浓度的3-甲基菲和2-甲基蒽处理健康仿刺参,检测其体内CYP450和p53基因的相对表达量,分析仿刺参CYP450和p53基因的表达与3-甲基菲和2-甲基蒽的剂量-效应关系和时间-毒性效应关系,为开展2种多环芳烃污染物对海洋生物毒性评价筛选敏感标志物。
1 材料与方法(Materials and methods)
1.1 仪器与试剂
仪器:Nano Photometer微量核酸蛋白分析仪(德国Implen),CT15RE型台式微量高速离心机(日本Hitachi),Mx3005pTM实时荧光定量PCR仪(美国Stratagene),ZHWY-200D恒温摇床(上海世平实验设备有限公司)。
试剂:2-甲基蒽和3-甲基菲购自Sigma公司(Sigma-Aldrich Corporation, USA),纯度分别大于97%和98%;丙酮(分析纯,上海国药集团化学试剂有限公司),Trizol RNA提取试剂盒(Invitrogen公司,上海),PrimeScriptTMRT reagent Kit(TaKaRa),SYBR PrimeScriptTMRT-PCR Kit II(TaKaRa)。
1.2 实验材料
实验用海水取自大连黑石礁海域的砂滤海水,仿刺参(A.japonicus)的幼参购自大连太平洋海珍品有限公司,体长(水中)为(2±0.3) cm,经清洁海水驯养2周后,选取健康幼参用于后续实验。
1.3 多环芳烃毒性实验
用1 L玻璃烧杯洗净后装入海水,加入一定浓度3-甲基菲和2-甲基蒽储备液,设置3-甲基菲浓度分别为5 μg·L-1、10 μg·L-1和100 μg·L-1,2-甲基蒽浓度分别为5 μg·L-1、10 μg·L-1和50 μg·L-1,同时设置海水对照组和丙酮对照组,各处理组设置3个平行,每个烧杯中放入10只幼参。试验条件:水温(15±2) ℃,盐度(30±1),pH (8.0±0.5),间断性充氧,确保溶解氧大于4.5 mg·L-1,避光。
1.4 样品采集及RNA提取
3-甲基菲和2-甲基蒽处理3 d、7 d、14 d后,取各处理组幼参一只放于冰上,用移液枪抽取海参体液,装入tube管中迅速投入准备好的液氮中,-80 ℃保存。Trizol法提取上述样品的总RNA,DNase I进行DNA消化处理,微量核酸蛋白分析仪检测总RNA的浓度和纯度,Agilent 2100生物分析仪检测RNA完整性。
1.5 基因表达水平测定
分别取上述样品总RNA用Prime-ScriptTMRT reagent Kit(TaKaRa)进行反转录。使用Mx3005pTM实时荧光定量PCR仪,采用SYBR Prime-ScriptTMRT-PCR Kit II进行实时定量PCR。以β-Actin作内参,相关基因扩增引物名称及其序列见表1。
反应条件:95 ℃,30 s;95 ℃,10 s;55 ℃,25 s;72 ℃,25 s,40个循环。利用delta-delta Ct法,通过仿刺参的内标基因AB510191.1(Actin)的校正,对仿刺参样品中CYP450和p53基因表达量进行分析。
1.6 数据统计与处理
试验数据用平均值±标准误差(mean±S.D.)表示,使用SPSS 22.0统计软件对所得数据进行统计学分析,采用单因素方差分析(One-Way ANOVA)和Duncan新复极差法对数据进行差异性分析。设置P<0.05时,表示差异显著。
2 结果(Results)
2.1 3-甲基菲、2-甲基蒽对仿刺参CYP450基因表达的影响
3-甲基菲和2-甲基蒽对仿刺参CYP450基因表达的影响如图1所示。结果表明,与对照组相比,3-甲基菲(c≥5 μg·L-1)和2-甲基蒽(c≥10 μg·L-1)在一定浓度胁迫下,对仿刺参CYP450基因的表达均表现出显著的抑制作用(P<0.05)。2-甲基蒽在浓度5 μg·L-1胁迫14 d后,对CYP450基因的表达有显著的诱导作用(P<0.05)。随着时间增加,海水对照组CYP450基因的相对表达量无明显变化,丙酮对照组CYP450基因的相对表达量不断增大,说明丙酮促进了仿刺参CYP450基因的表达。与对照组相比,3-甲基菲暴露14 d后,CYP450基因的整体相对表达量小于对照组,差异显著(P<0.05),随着浓度的升高,CYP450基因的相对表达量逐渐增大,100 μg·L-1处理组与10 μg·L-1处理组相比,差异不显著(P>0.05)。用2-甲基蒽暴露14 d后,随着浓度的升高CYP450基因的表达量受到明显的抑制,且存在一定的剂量-效应关系(P<0.05)。连续暴露14 d后,仿刺参CYP450基因相对表达量在3-甲基菲浓度为5 μg·L-1胁迫下达到最小值0.21,在2-甲基蒽浓度为50 μg·L-1胁迫下达到最小值0.16。
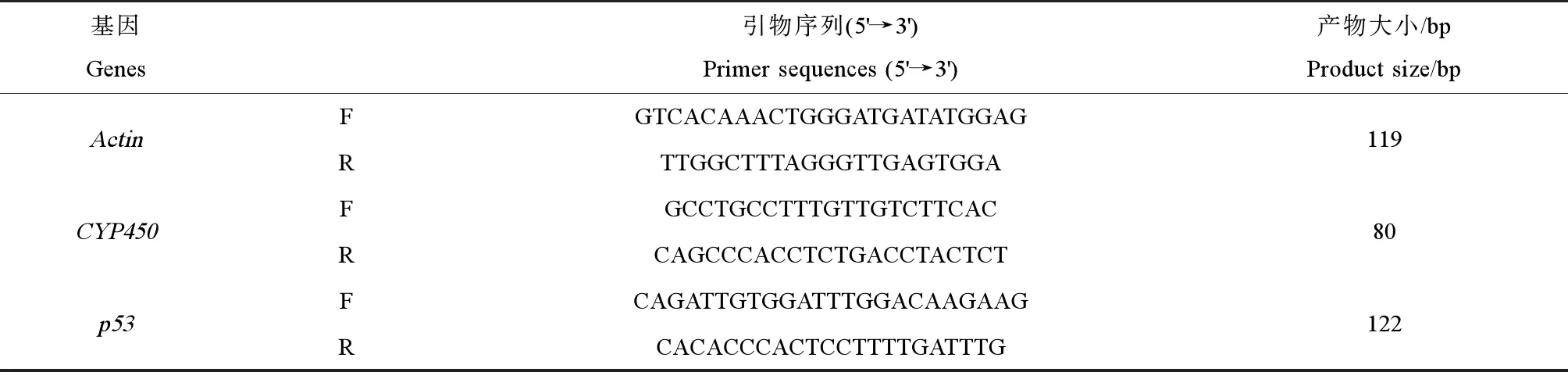
表1 相关基因扩增引物名称及其序列Table 1 Name and sequence of the target gene amplified with primers

图1 3-甲基菲、2-甲基蒽对仿刺参CYP450基因表达的影响注:标有不同小写字母者表示显著性差异(P<0.05),标有相同小写字母者表示无显著性差异(P>0.05),图表横坐标中B表示空白对照组,B-B表示溶剂对照组,下同。Fig. 1 The effects of 3-methylphenanthrene, 2-methylanthracene on the CYP450 gene expression of Apostichopus japonicusNote: The different letters mean significant differences at the 0.05 probability level, and the same letters mean no significant differences; B represents the seawater control group and B-B represents the acetone control group.
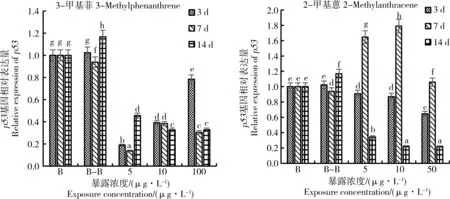
图2 3-甲基菲、2-甲基蒽对仿刺参p53基因表达的影响Fig. 2 The effects of 3-methylphenanthrene, 2-methylanthracene on the p53 gene expression of Apostichopus japonicus
2.2 3-甲基菲、2-甲基蒽对仿刺参p53基因表达的影响
3-甲基菲和2-甲基蒽仿刺参p53基因表达的影响如图2所示。3-甲基菲和2-甲基蒽胁迫初期(3 d)对仿刺参p53基因的相对表达量表现出规律不一致的变化,与对照组相比,3-甲基菲各处理组对仿刺参p53基因的表达均表现出显著的抑制作用(P<0.05),且随着胁迫浓度的升高,抑制作用逐渐减小,存在明显的剂量-效应关系,2-甲基蒽各处理组对仿刺参p53基因的表达也均表现出显著的抑制作用(P<0.05),但随着胁迫浓度的升高,抑制作用逐渐增大。3-甲基菲胁迫中期(7 d),随着胁迫浓度的升高,仿刺参p53基因的相对表达量表现出先升高后降低的规律,2-甲基蒽胁迫中期(7 d),各处理组对仿刺参p53基因的表达均表现出诱导作用,且随着胁迫浓度的升高,仿刺参p53基因的相对表达量表现出与3-甲基菲胁迫相似的规律,也呈先升高后降低的趋势。3-甲基菲和2-甲基蒽胁迫后期(14 d),与对照组相比,各处理组均对仿刺参p53基因的表达表现出显著的抑制作用(P<0.05),且均在胁迫浓度为10 μg·L-1时仿刺参p53基因的相对表达量达到最小值。
3 讨论(Discussion)
CYP450和p53基因的表达在3-甲基菲和2-甲基蒽污染物胁迫下整体上均表现出抑制作用,说明在外源污染物进入仿刺参体内时,CYP450和p53基因的表达均能对毒物产生响应以维持仿刺参正常的生理活动,但二者的具体响应程度及对2种污染物的耐受阈值存在差异,这可能与CYP450基因和p53基因在生物体内的主要功能及作用机制不同有关。不同的外源化学物质进入生物体内后可引起氧化应激、基因毒性应激和蛋白质毒性应激等反应,从而激活生物细胞内解毒代谢系统,以维持生物体内环境的相对稳定[14]。而检测参与这些反应的各种解毒代谢酶类的响应变化可部分反映外源污染物对生物的影响。环境污染物进入生物体内后,通常会在P450酶系的参与下,进行氧化、还原、水解等代谢过程,使得污染物极性增加,导致污染物的毒性减弱[15]。海洋无脊椎动物在有机污染物的I相代谢过程中产生大量副产物——活性氧,过量的活性氧会导致生物体的氧化损伤[16-19]。CYP450酶系作为生物体内的第一代谢阶段酶,它通常将分子氧拆分成原子氧,并将其加入到底物中促使底物羟基化或环氧化,为第二阶段的谷胱甘肽转移酶(GST)对外源物的转化提供基础[20]。在本实验中,仿刺参CYP450基因的表达在3-甲基菲胁迫时受到抑制,在2-甲基蒽胁迫3 d和7 d后均受到抑制,而胁迫14 d后表现出显著的诱导作用(P<0.05)。因此,仿刺参CYP450基因的表达在不同多环芳烃胁迫下表现出不一致的变化规律。这与菲和3-甲基菲等对河鲀(Takifugurubripes)幼鱼肝组织损伤的影响及多环芳烃类物质对CYP450 1A1的诱导表达表现出相似的规律[2,21]。因此,推测短时间内(7 d)2-甲基蒽诱导仿刺参CYP450基因的表达,调动相关酶来抵抗毒性物质的毒性作用,降低或减轻细胞的应激反应,但其调节能力有限,CYP450基因的表达不可能无限提高,当随着染毒时间的延长(14 d),仿刺参受到的毒性作用超出其耐受性,其CYP450的合成体系受到损伤,使得CYP450基因的相对表达量下降,表现出中毒效应。而CYP450基因对3-甲基菲的耐受能力较强,在染毒期间持续表现为诱导作用。因此,多环芳烃类污染物3-甲基菲对仿刺参CYP450基因表达的影响小于2-甲基蒽。
p53基因是化学诱变物的主要攻击位点,p53基因突变后,失去了对细胞生长、凋亡和DNA修复的调控作用。p53不仅本身参与机体DNA损伤的修复过程,它还作为多种细胞应激于细胞应答的中间环节,与其上下游的各种调控因子及其相关基因共同构成了调控细胞应激和细胞应答的复杂网络[22]。在本实验中,仿刺参p53基因的表达在3-甲基菲胁迫时受到了抑制,在2-甲基蒽胁迫7 d时表现出显著的诱导作用(P<0.05),而后(14 d)表现出显著的抑制作用。正常组织中p53的表达水平很低,当生物受到外界胁迫时,p53的表达水平明显增加[23]。有研究报道,纳米二氧化硅包被氧化锰能诱导Hela细胞和L929细胞中p53基因表达量升高[24]。因此推测,3-甲基菲在低浓度(c≤100 μg·L-1)短时间(t≤14 d)胁迫时没有对仿刺参造成较强的毒害作用,而2-甲基蒽胁迫长时间(7 d)时对仿刺参产生严重的毒害作用,且仿刺参p53基因对2-甲基蒽胁迫时间有一定的耐受阈值,当胁迫时间大于此阈值后,可能导致细胞损伤或死亡,致使p53基因相对表达量减少。
比较2种多环芳烃类污染物对仿刺参CYP450和p53基因的相对表达量发现,在相同浓度和时间胁迫下,2种物质对仿刺参CYP450和p53基因表达的影响作用总体趋势为3-甲基菲<2-甲基蒽,这可能意味着由于二者结构和性质的差异导致其作用机制不同。除浓度和时间外,化合物的结构可能对其毒性有重要影响,需进一步进行相关研究。
致谢:感谢大连海洋大学徐光景博士在文章修改中给予的帮助。
猜你喜欢
分子催化(2022年1期)2022-11-02
水泵技术(2022年3期)2022-08-26
化学工业与工程(2022年1期)2022-03-29
红蜻蜓·低年级(2021年12期)2022-01-19
红蜻蜓·低年级(2021年12期)2021-12-19
石油沥青(2021年4期)2021-10-14
渔业科学进展(2021年3期)2021-05-12
科学家(2021年24期)2021-04-25
大连海洋大学学报(2020年2期)2020-05-06
中成药(2019年12期)2020-01-04
