
槐耳清膏对胃癌细胞NCI-N87自噬的影响*
2019-01-08 03:05廖传文李洁兰殷俊翔胡淑琴饶雪峰
江西医药 2018年11期
廖传文 ,李洁兰 ,殷俊翔 ,胡淑琴 ,饶雪峰
(1.江西省人民医院普外科,南昌 330006;2.江西省吉水县人民医院,吉水 331600)
中药在抗肿瘤方面扮演着非常重要的作用[1],槐耳提取物在中国有1600年的使用历史[2],研究表明槐耳提取物在体外可抑制多种肿瘤细胞活性及增殖作用[3]。自噬是一个保守的依赖溶酶体的降解途径,可降解长寿命蛋白质、细胞器和部分细胞质,并已被证明参与包括能量代谢、细胞器周转、生长调节和衰老等生理病理过程[4,5]。近年来研究表明,自噬在肿瘤细胞生长、分化和抵抗药物治疗等方面扮演重要角色[6]。槐耳是否能通过影响胃癌细胞自噬进而影响胃癌的发生发展,尚鲜有报道。本研究以胃癌细胞NCI-N87为研究对象,探讨槐耳清膏对NCI-N87细胞自噬影响及可能的机制。
1 材料与方法
1.1 材料 槐耳清膏 (江苏启东盖天力公司),DMEM溶解后,0.22um滤膜过滤,加入10%PBS,使其终浓度分别为 4、8、16、32mg/ml。 胃癌细胞株NCI-N87购于中国生命科学院上海细胞库。啶橙购 自 Sigma 公 司 ,Beclin 1、PTEN、LC3-II 和GAPDH抗体购于Santa cruz公司;TRIzol试剂购于invitrogen公司;逆转录试剂盒为Promega公司产品;Real-Time PCR试剂盒为fermentas公司产品;Real-Time PCR扩增仪为美国ABI公司产品;流式细胞仪为美国BD公司产品;引物由上海生工生物技术公司合成。半干转印系统转印槽为Bio.Rad公司产品。
1.2 方法
1.2.1 细胞培养 NCI-N87细胞株常规在含10%胎牛血清的0.1%DMSO培养基,于37℃、5%CO2的孵箱中培养,用0.25%胰蛋白酶消化,在显微镜下观察细胞形态,待细胞长满至80%融合度时,以1:3比例进行传代。使活细胞数>95%,取对数生长期的细胞用于实验。
1.2.2 CCK-8法检测细胞抑制率 将对数生长期NCI-N87细胞接种于96孔培养板中,每孔细胞接种数目为2000个,同时每孔加入培养液100μl,另设不加细胞的培养液为本底,24h后实验组分别加入槐耳清膏10μl,使其最终浓度分别为4、8、16、32mg/ml。空白对照组为未加药物的细胞,加入等量培养液。每个浓度设有6个复孔,然后在37℃、5%CO2培养箱中继续培养24、48和72h,每日相同时间点将不同浓度组的细胞放置于倒置显微镜下观察其细胞形态改变,并于时间点向各个孔培养液中分别加入CCK-8试剂各10μl,然后于5%CO2、37℃环境中继续培养4h后,使用酶标仪测试吸光度,测定标准为490nm的的吸光度波长,记录所有孔的吸光度(OD)的数值,计算细胞抑制率(%)。细胞抑制率 (%)=[1-(实验组OD值一空白组OD值)/(对照组OD值一空白组OD值)]×100%。
1.2.3 AnnexinV/PI流式细胞仪检测细胞凋亡 将对数生长期的NCI-N87细胞制成悬液,按5×103个/ml接种于6孔板中,共设6组,每组设3个复孔,向各实验组分别加入含浓度为4、8、16、32mg/ml槐耳培养液,处理24h。对照组加入含0.1%DMSO培养基。检测前用PBS洗涤细胞2次,按照说明书进行以下操作:每管加入5μl的AnnexinV和PI复合染液和500μl结合缓冲液,避光染色20min后,进行流式细胞仪检测,分析槐耳处理后的细胞凋亡百分比变化。
1.2.4 细胞自噬小体的定量检测 不同浓度槐耳清膏(4、8、16、32mg/ml)作用 NCI-N87 细胞 24h 后,收集细胞,用浓度为1μg/ml吖啶橙室温条件下避光染色20min,离心之后弃染液,用1×PBS清洗3次后,PBS重悬细胞,上流式细胞仪检测,FL3通道检测红色荧光,FL1通道检测绿色荧光,红色荧光增强说明细胞内酸性自噬小体增多。
1.2.5 RT-PCR 检测 Beclin-1、LC3-II、PTENmRNA表达 分别检测不同浓度的槐耳作用NCI-N87细胞24h后,按照说明书操作,用Trizol试剂提取各组细胞总RNA,紫外分光光度仪测定RNA浓度,并行纯度验证、电泳鉴定,然后进行逆转录,以cDNA为模板进行Real-Time PCR反应。PCR产物以琼脂糖凝胶电泳进行分离,凝胶成像分析系统行密度分析。GAPDH作为内参照。目的基因mRNA表达量以各目的基因片段密度值/内参照密度值的比值表示。Beclinl基因上游引物5’-ATCCTGGACCGTGTCACCATCCAGG-3’, 下游引物 5’-GTTGAGCTGAGTGTCCAGCTGG-3’。LC3-II上游引物 5’-ACCATGCCGTCGGAGAAG3’; 下游引 物 5’ -ATCGTTCTATTATCACCGGGATTT-3’。PTEN上游引物 5’-TCACCAACTGAAGTGCCTAA AGA-3’;下游引物 5’-CTCCATTCCCCTAACCCGA-3’。 GAPDH 上游引物 5’-GTAAAGACCTCTATG CCATCA-3’;下游引物 5’-GGACTCATCGTACTC CTGCT-3’。
1.2.6 Westernblot检测 Beclin-1、LC3-II、PTEN 蛋白表达 取对数生长期的NCI-N87细胞,并用不同浓度槐耳清膏(8、16、32mg/ml)处理细胞 24h;实验前2h用含有2%胎牛血清的RPMI-1640预处理细胞,收集处理后的细胞,加入裂解液,冰上裂解30min后,超声破碎,4℃、离心10min,转速15000r/min,取上清,采用BCA法测定蛋白质含量,SDSPAGE凝胶电泳,低温转移至PVDF膜,封闭,分别加入 Beclin-1、LC3-II、PTEN 一抗 4℃过夜、山羊抗兔二抗30℃温育1h后显色,用数字成像仪对图像进行拍照和分析。上述实验重复3次。
2 结果
2.1 槐耳清膏对NCI-N87细胞增殖影响 槐耳清膏(4、8、16、32mg/ml)作用 NCI-N87 细胞 24、48、72h后, 与对照组相比,8、16、32mg/ml对胃癌细胞有抑制增殖作用,并以浓度和时间依赖的方式抑制细胞增殖(P<0.05),而低浓度组 4mg/ml未显现明显抑制作用。结果见表1。
表1 不同浓度槐耳清膏作用不同时间后对胃癌细胞NCI-N87存活的影响()

表1 不同浓度槐耳清膏作用不同时间后对胃癌细胞NCI-N87存活的影响()
注:与对照组比较,★P<0.05;与同组 24h 比较,◆P<0.05;与同组 48h比较,#P<0.05。
1.02±0.19 0.88±0.18 0.51±0.08★◆#0.42±0.06★◆#0.32±0.02★◆#组别对照组4mg/ml 8mg/ml 16mg/ml 32mg/ml 24h 1.06±0.23 0.95±0.18 0.78±0.17★0.63±0.19★0.42±0.04★48h 72h 1.04±0.41 0.89±0.19 0.64±0.17★◆0.51±0.17★◆0.39±0.09★◆
2.2 槐耳清膏对NCI-N87细胞凋亡率及自噬小体的影响 槐耳清膏(8、16、32mg/ml)作用 NCI-N87细胞24h后,流式细胞分析结果显示,与对照组相比,8、16、32mg/ml浓度能促进 NCI-N87 细胞凋亡,自噬小体形比例增加。(P<0.05)。结果见表2。
表2 槐耳清膏对胃癌细胞NCI-N87凋亡率及自噬小体的影响(,%)
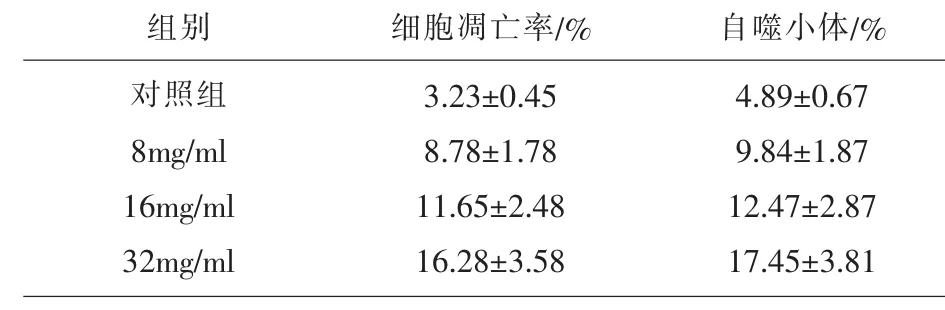
表2 槐耳清膏对胃癌细胞NCI-N87凋亡率及自噬小体的影响(,%)
注:与对照组比较,★P<0.05。
自噬小体/%4.89±0.67 9.84±1.87 12.47±2.87 17.45±3.81组别对照组8mg/ml 16mg/ml 32mg/ml细胞凋亡率/%3.23±0.45 8.78±1.78 11.65±2.48 16.28±3.58
2.3 槐耳清膏对NCI-N87细胞相关自噬蛋白表达的影响 通过RT-PCR和Western blot对自噬相关蛋白Beclin-1、LC3-II、PTEN mRNA和蛋白水平进行了检测,结果表明,与对照组相比,槐耳清膏作用 NCI-N87 细胞 24h 后,Beclin-1、LC3-II、PTEN mRNA水平和蛋白表达水平逐渐升高,均有统计学意义(P<0.05)。 结果见图 1、图 2、图 3 及图 4。
3 讨论
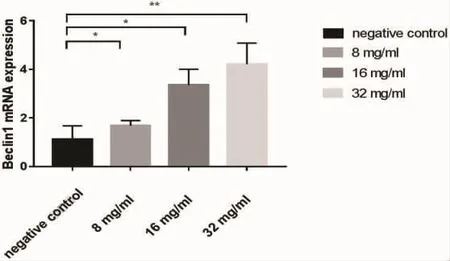
图1 不同浓度槐耳清膏对NCI-N87细胞Beclin-1mRNA表达水平影响

图2 不同浓度槐耳清膏对NCI-N87细胞LC3-II mRNA表达水平影响

图3 不同浓度槐耳清膏对NCI-N87细胞PTEN mRNA表达水平影响
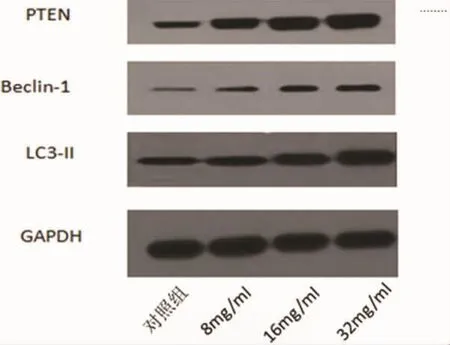
图4 槐耳清膏对NCI-N87细胞Beclin-1、LC3-II、PTEN蛋白表达影响
胃癌是消化道最常见的恶性肿瘤之一,在因肿瘤而死亡的患者中,胃癌占第二位[7,8]。HER2在胃癌中的阳性表达率为3.9%-51.1%,HER2的过表达与胃癌的Bormann分型、Lauren分型、肿瘤的分化程度、淋巴结的状态、静脉清润等有关,而与肿瘤的大小、清润的深度及肿瘤的分期无关[9]。
自噬不仅参与了正常细胞生长发育、同时也参与了细胞的成熟分化及死亡的调控,自噬活性的改变经常可见于一些肿瘤细胞,影响了肿瘤的发生和发展[4,6]。
大量研究表明,槐耳具有抗肿瘤活性,可以抑制血管及肿瘤形成、抗肿瘤耐药、抑制癌转移、激活免疫系统以及诱导凋亡等[10,11]。最近也有研究报告,槐耳清膏能诱导乳腺癌细胞发生自噬及凋亡,其机制可能是通过抑制MTOR/S6K通路有关[12],另有研究结果表明,槐耳清膏对胃癌MKN-45细胞增值及凋亡均有影响[13],但能否影响her-2阳性胃癌细胞自噬尚未见相关报道。
在本研究中,我们以高表达her-2的NCI-N87胃癌细胞株为研究对象,通过研究不同浓度的槐耳清膏在处理细胞不同时间后,对NCI-N87细胞增殖、凋亡及相关自噬蛋白表达的影响。
研究中我们首先观察到,大于4mg/ml浓度的槐耳清膏对于细胞的增殖具有明显的影响,在我们观察的24、48、72h内,随着时间延长其抑制作用增加,呈现质量浓度和时间的依赖关系。胞质内大量的酸性自噬小体形成是自噬的重要特征,本实验通过流式细胞仪检测发现细胞内自噬小体和细胞凋亡率均增加,且与浓度相关。上述的研究结果表明,槐耳清膏能影响NCI-N87增殖、凋亡及自噬,从而能在体外影响NCI-N87细胞,发挥抑癌作用。
有研究表明,细胞自噬受到自噬基因的调控,如 Beclin-1、LC3-II、PTEN。
有研究报道自噬相关蛋白Beclin-1在胃癌组织和正常组织中出现表达差异,正常组织阳性表达高于胃癌组织,是与自噬相关的抑癌基因,它作为激活自噬信号通路的关键蛋白之一,可以调控凋亡信号通路,从而诱导癌细胞的凋亡[14]。并且进一步的研究表明Beclin-1的蛋白表达水平也与胃癌组织的分化程度相关[15,16]。另外,自噬体的形成需要多种蛋白质复合体和小分子的参与,其标志性蛋白是LC3,LC3介导的自噬作用在胃癌中至关重要[17]。LC3共分为 LC3-I和 LC3-II两型,其LC3-I和LC3-II的含量和比值变化能够反映细胞的自噬活性[18]。在恶性程度高的胃癌患者中,Beclin-1表达呈现逐渐降低的趋势,而LC3-II的蛋白表达则呈现逐渐上升的趋势,两者均与胃癌的发生发展密切相关[19]。而PTEN基因作为一个具有脂性磷酸酯酶活性的抑癌基因,可通过PI3KAKT-mTOR通路和PI3K/AKT通路抑制脂性磷酸酶活性,从而促进细胞的自噬过程。因此,PTEN蛋白被认为是诱导自噬发生的正向调节分子,这也提示了PTEN通过自噬对抑制胃癌的发生发展具有重大意义[20,21]。
在本研究中,我们发现大于4mg/ml浓度的槐耳清膏作用 NCI-N87 24h 后,Beclin-1、LC3-II、PTEN在mRNA水平及蛋白水平均明显升高,结果具有统计学意义,且与浓度呈现正相关性。从上述结果我们推测,槐耳清膏可能影响自噬基因Beclin-1表达,同时增加PTEN表达,导致自噬发生,较高浓度的槐耳清膏效果明显,但超过32mg/ml是否自噬效应增加尚没有试验结果证明。
综上所述,槐耳清膏能够在体外促进NCIN87自噬和凋亡,同时能够抑制NCI-N87细胞增殖,在一定的浓度及作用时间范围内两者呈正相关关系。其可能的机制是槐耳清膏可能影响自噬基因Beclin-1表达,同时增加PTEN表达,导致自噬发生。由于自噬的机制甚为复杂,在本研究中槐耳清膏是否影响自噬相关信号通路改变,尚不清楚,能否在体内发生抑瘤作用也是我们进一步研究的方向。
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
中西医结合心脑血管病杂志(2022年19期)2022-11-19
检验医学与临床(2022年12期)2022-06-27
中国人兽共患病学报(2022年5期)2022-06-08
江西医药(2018年8期)2018-10-24
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
中国卫生标准管理(2015年3期)2016-01-14
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年20期)2015-03-01
