
miR-133调控FGFR1蛋白对喉癌细胞迁移和侵袭的影响
2018-12-26 09:03潘庆春喻望博李四军
解放军医药杂志 2018年12期
潘庆春,喻望博,李四军
喉癌是全球常见的头颈部恶性肿瘤,而鳞状细胞癌是最常见的一种病理类型[1-2]。然而,喉癌被诊断时通常处于中晚期阶段,并常伴有局部病灶浸润扩散和远处淋巴结的转移[3]。尽管随着诊断技术的进步和治疗方式的改善,总体的生存率有所提升,但仍然不尽如人意[4]。现在学者们更注重对肿瘤分子特征进行区别,并根据特定的基因组差异进行肿瘤的亚型分类[5]。根据精确的肿瘤分类所开发出的针对精确分子改变的临床治疗称为靶向治疗[6-7]。这种治疗方法为癌症患者提供了更为有效的治疗,并且与常规化学疗法相比,通常具有更小的毒副作用[8]。miRNAs是一类非编码RNA,能通过与编码蛋白RNA的3' -UTR端结合,从而影响基因的功能与表达[9]。miRNA通过结合与之相同或相似的互补序列来调节基因表达,引起翻译的抑制或切割mRNA靶标[10]。miR-133已被证实在多种癌症的发生发展过程中起到重要作用[11-15],能抑制肿瘤细胞的增殖、迁移、侵袭和细胞周期进展[16]。但miR-133在喉癌发生发展过程中的作用尚不清楚。本研究拟探讨miR-133对喉癌细胞的影响及其相关作用机制。
1 材料与方法
1.1组织标本 选取2017年1月—2018年1月我院收治20例喉癌患者的肿瘤组织及癌旁正常组织。患者年龄37~68岁,平均52岁;病例样本病理类型均为鳞状细胞癌,临床病理分期均为Ⅰ~Ⅱ期。将取下的组织样品在液氮中快速冷冻。所有患者手术前没有接受过放疗或化疗。本研究经医院伦理委员会批准所有实验组织标本的使用。
1.2细胞培养 人喉癌细胞株Hep-2和TU212购买于武汉大学;RPMI-1640培养基和0.25胰酶(0.25 Trypsin EDTA)购买于美国Gibco公司;胎牛血清和双抗(Penicillin-Streptomycin Solution)购买于美国海克隆公司;PBS磷酸盐缓冲液购买于BOSTER。细胞均培养在含10%胎牛血清和1%双抗的培养基中,所有细胞均在5%CO2,37℃条件下培养,每隔3 d换液1次,待细胞铺满培养瓶底进行传代,以下所有试验均取处于对数生长期的细胞。
1.3细胞转染 miR-133 mimics和miR-133 inhibitor购买于上海吉玛有限公司。PBS缓冲液清洗Hep-2和TU212细胞,应用胰酶消化2 min,转移至无菌15 ml离心管,离心并计数细胞,以每孔4×105个细胞接种于6孔板中,使过夜细胞融合率达70%左右,细胞分为Hep-2 mimics组、Hep-2 NC组、Hep-2 inhibitor组、TU212 mimics和TU212 NC组,其中mimics组细胞均转染miR-133 mimics慢病毒,inhibitor组细胞均转染miR-133 inhibitor慢病毒,NC组细胞均转染NC慢病毒。无血清培养基将Lipofectamine 2000按3 μl/L稀释,37℃孵育20 min,用无血清培养基分别将miR-133 mimics和miR-133 inhibitor按50 μmol/L浓度稀释,常温下孵育5 min,与Lipofectamine 2000等体积分别混匀,分别加入细胞中,37℃孵育箱继续培养;12 h后,观察转染细胞状态,并将无血清培养基更换为完全培养基,继续培养48 h后,提取细胞RNA,验证转染效率。
1.4定量实时PCR分析 氯仿、乙醇、异丙醇购买于重庆川东化工公司;Trizol裂解液、Premix ExTaq Version2.0和SYBR Premix Ex Taq购买于日本Takara公司。PCR引物由上海Sangon Biotech公司合成。miR-133的qRT-PCR检测以U6为内参,采用2-ΔΔCt方法进行统计,重复3次,利用荧光PCR仪检测cDNA中miR-133的相对表达量,反应条件为:95℃预变性10 min,95℃ 15 s,60℃ 15 s,45个循环,获取荧光信号温度为60℃。普通cDNA的qRT-PCR检测以GAPDH为内参,采用2-ΔΔCt方法进行统计,反应条件为95℃ 10 min,95℃ 15 s,60℃ 1 min,共40个循环。
1.5划痕愈合实验 将喉癌细胞以每孔2×104个细胞密度接种于6孔板,融合度达到后,用200 μl消毒枪头垂直划线,并于显微镜下测量划痕的初始距离,在恒温孵箱中孵育72 h后,再次测量划痕的距离,并计算细胞的迁移率。实验重复3次。
1.6Transwell侵袭试验 Transwell基质胶购买于美国BD公司;Transwell小室购买于美国康宁公司。将5×104个细胞置于上室,并加入Matrigel基质胶。将含10%胎牛血清和HGF(20 μg/ml)的培养基置于下室。37℃孵育24 h,去除上膜表面的细胞。95%乙醇固定20 min,0.5%结晶紫溶液染色10 min,自来水冲洗后,在倒置显微镜下计数。
1.7荧光素酶报告基因测定 构建含成纤维细胞生长因子受体1(FGFR1)基因野生型3'-UTR(含miR-133识别位点/wt)和突变型3'-UTR(miR-133识别位点点突变/mut)片段,两端均设计XbaⅠ酶切位点,插入荧光素酶报告基因质粒构建pGL3-3'-UTR wt、pGL3-3'-UTR mut。取生长状态良好的喉癌Hep-2细胞,质粒转染后48 h据操作说明,裂解细胞,加入荧光素酶底物,检测荧光素酶活性,以证明miR-133直接调控PRDX1。
1.8Western blotting实验 GAPDH小鼠抗人一抗、β-actin小鼠抗人一抗购买于武汉三鹰生物技术有限公司;Western blotting凝胶试剂盒和全蛋白提取试剂盒购买于碧云天公司;ECL发光液购买于索莱宝生物科技有限公司。根据蛋白分子量大小,配制凝胶,根据所测蛋白浓度加入蛋白样品以及每孔5 μl蛋白marker;跑胶,首先加压80 V,保证所有样品在同一水平线,待样品至分离胶处,将电压加到120 V;切胶,根据所需条带大小,对照蛋白marker进行切胶,并准备与所切胶条相同大小滤纸2片和PVDF膜(浸泡甲醇15 s);按照滤纸、胶、PVDF膜、滤纸的顺序按序排好,并赶出气泡,压紧,在250 mA恒流下,按1 kD蛋白1 min进行转膜;转膜结束后,PBST洗膜1次,脱脂奶粉封闭1 h;PBST洗膜1次,将膜浸入稀释好的一抗中,4℃过夜;PBST洗膜3次,将膜浸入稀释好的二抗中,常温下孵育1 h;显影,PBST洗膜3次后,在暗室中曝光,并分析。
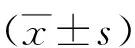
2 结果
2.1miR-133的表达情况 miR-133在喉癌组织和癌旁正常组织的表达分别为0.57±0.12和1.09±0.02。与癌旁正常组织比较,喉癌组织中miR-133呈显著低表达状态(P<0.05)。
2.2miR-133对Hep-2和TU212细胞迁移的影响 Hep-2 mimics组和Hep-2 NC组的细胞的迁移率分别为(40.1±10.4)%和(98.4±9.5)%,TU212 mimics组和TU212 NC组的细胞的迁移率分别为(58.6±8.0)%和(99.2±10.1)%。与Hep-2 NC组和TU212 NC组比较,Hep-2 mimics组和TU212 mimics组的细胞迁移率均显著降低(P<0.05)。见图1。
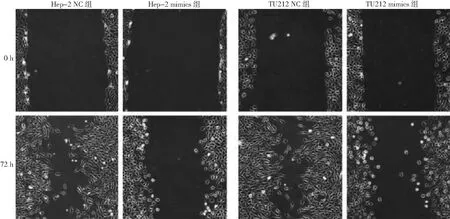
图1 miR-133对人喉癌Hep-2和TU212细胞迁移的影响
2.3miR-133对Hep-2和TU212细胞侵袭的影响 Hep-2 NC组和Hep-2 mimics组细胞侵袭率分别为(98.4±7.5)%和(48.1±10.4)%,TU212 NC组和TU212 mimics组细胞侵袭率分别为(97.2±10.1)%和(38.2±5.3)%。与Hep-2 NC组和TU212 NC组比较,Hep-2 mimics组和TU212 mimics组的细胞侵袭率显著降低(P<0.05)。见图2。

图2 miR-133对人喉癌Hep-2和TU212细胞侵袭的影响
2.4miR-133靶向调控FGFR1 通过Targetscan网站发现miR-133与FGFR1具有相似的结合位点。见图3。通过双荧光素酶报告基因验证二者的关系,结果显示野生型和突变型FGFR1基因的荧光素酶活性分别为0.26±0.02和0.96±0.06,比较差异有统计学意义(P<0.05)。表明miR-133可以明显抑制野生型FGFR1基因的荧光素酶活性,而对突变型FGFR1基因的荧光素酶活性无影响。Western blotting结果也表明,Hep-2 mimics组、Hep-2 inhibitor组和Hep-2 NC组FGFR1蛋白表达分别为(31.8±8.0)%、(96.1±9.6)%和(73.2±6.8)%,3组比较差异有统计学意义(P<0.05)。miR-133的过表达和沉默能使Hep-2细胞中的FGFR1蛋白表达量下调和上调。见图4。

图3 FGFR1基因与miR-133的3'-UTR基因序列比对FGFR1为纤维细胞生长因子受体1

图4 miR-133对Hep-2细胞FGFR1蛋白表达的影响FGFR1为纤维细胞生长因子受体1
3 结论
喉癌患者存在miRNAs的失调,有研究显示,miRNAs在喉癌的增殖、转移、侵袭和凋亡过程中发挥重要作用[17]。miRNAs在喉癌中可以发挥致癌作用或抑癌作用[18]。Cao等[19]通过研究发现,共有26个miRNAs在喉鳞状细胞癌组织中高表达,有3个miRNAs显著低表达,而且还发现,miR-21、miR-93、miR-205和miR-708是显著上调的,而miR-125b和miR-145显著下调。本研究结果发现,喉癌组织中miR-133呈显著低表达状态。表明miR-133在喉癌中可能是扮演抑癌基因的角色。
不同miRNAs对喉癌细胞的影响可能通过不同的行为方式及相应的机制。如miR-1通过直接影响纤维连接蛋白1的途径抑制Hep-2细胞的生长、迁移和侵袭[20]。Guo等[21]研究发现,miR-24通过下调S100A8诱导Hep-2细胞形态改变并抑制细胞增殖和侵袭。另外,miR-30b参与调节重要的肿瘤抑制基因p53,在miR-30b过表达的Hep-2细胞中p53表达显著增强,同时miR-30b过表达增加p53介导的肿瘤细胞凋亡[22]。本研究结果发现,与Hep-2 NC组和TU212 NC组比较,Hep-2 mimics组和TU212 mimics组细胞的侵袭率和迁移率显著降低。表明miR-133的过表达能显著抑制Hep-2和TU212细胞的增殖和迁移能力。
成纤维细胞生长因子受体(FGFRs)是一种酪氨酸激酶,位于染色体8p12上,与多种肿瘤侵袭行为密切相关,而且能一定程度上预测患者的生存和预后[23-24]。FGFRs家族包括酪氨酸激酶的亚组受体,由5个结构保守的反式膜糖蛋白组成,含有3个免疫球蛋白样结构域在其胞外区[25]。被认为是真正的致癌基因,可能在肿瘤发展过程中发挥关键作用[26]。FGFRs及其配体,即成纤维细胞生长因子(FGF),参与多种正常生物学进程,包括细胞分化、增殖、迁移和血管生成等[27]。大量证据还表明,FGF-FGFR途径在肿瘤发展过程中起关键作用,并可能促进上皮间质转化,并激活关键的致癌信号通路,包括PI3K/AKT,RAS/MAPK和TGFβ途径[28]。
FGFR1的高表达也在头颈部的恶性肿瘤中被发现,而且其上调多见于人乳头状瘤病毒阴性鳞癌[29]。与肺癌类似,FGFR1的升高预示着喉癌患者生存率的降低[30]。目前,对喉癌尚无针对FGFR1的靶向治疗。本研究通过Targetscan网站发现,miR-133与FGFR1具有相似的结合位点,两者可能有内在调控关联。本研究还发现,miR-133可以明显抑制野生型FGFR1基因的荧光素酶活性,而对突变型FGFR1基因的荧光素酶活性无影响。表明miR-133可以与FGFR1的3'-UTR特异性结合。同时miR-133的过表达和沉默能使Hep-2细胞中的FGFR1蛋白表达量下调和上调。以上结果均证实miR-133能靶向调控FGFR1的表达,从而进一步影响肿瘤细胞恶性生物学进程。
综上所述,miR-133能通过靶向调控FGFR1蛋白的表达从而影响喉癌细胞迁移和侵袭能力。miR-133可能会成为治疗喉癌的新的靶点。
猜你喜欢
中风与神经疾病杂志(2021年9期)2021-11-08
天津医科大学学报(2021年4期)2021-08-21
婚育与健康(2021年5期)2021-07-06
中日友好医院学报(2021年1期)2021-04-14
山东医药(2020年9期)2020-05-20
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
标记免疫分析与临床(2016年9期)2016-11-21
中国当代医药(2015年1期)2015-03-01
中国医药导报(2015年27期)2015-02-28
