
ARTP诱变选育亚油酸异构酶高产菌株及其发酵条件优化
2018-12-21 06:49:58崔晓峰薛正莲
徐州工程学院学报(自然科学版) 2018年4期
张 珂,魏 明,崔晓峰,薛正莲
(安徽工程大学 生物与化学工程学院,安徽 芜湖 241000)
共轭亚油酸(CLA)是亚油酸在空间位置和几何构型上的一系列同分异构体的总称[1],具有抗肿瘤、抗氧化、防治糖尿病等功能[2-4],在新型药品、营养添加剂以及功能性食品等领域有重要的应用价值[5-6].生物法合成CLA特异性高,条件温和,但转化率较低[7].因此,对亚油酸异构酶高产菌株的诱变选育及其发酵条件的优化一直是人们工作的重点.
迄今,紫外辐照[8]、亚硝基胍诱变[9]等方法分别被用于油酸异构酶高产菌株的诱变选育,不同程度提升了CLA的产量.常压室温等离子体诱变(ARTP)技术是一种新型的微生物基因组快速诱变技术,具有诱变环境简单,诱变温度低,操作便捷等优点.而对突变株发酵条件的优化一直是人们研究提高CLA产量的基础.本研究以ARTP技术对油酸异构酶产生菌进行多轮诱变选育,并通过单因素及多因素响应面实验对高产突变株的产酶条件进行优化,以获得最佳发酵条件,为生物法生产CLA提供理论基础.
1 材料与方法
1.1 材料与仪器
嗜酸乳杆菌(L.acidophilus)CICC6075,购自中国工业微生物菌种保藏管理中心;共轭亚油酸标准品、溶菌酶,Sigma公司;亚油酸(纯度>95%)、正己烷、吐温-80、葡萄糖等常规试剂均为分析纯,购于国药集团化学试剂有限公司.
pH计、分析天平,梅特勒-托利多国际股份有限公司;ARTP育种,无锡思清源生物科技有限公司;紫外可见分光光度计,上海光谱仪器有限公司;超声波细胞粉碎仪,宁波新芝生物科技股份有限公司;全自动高压灭菌器,日本Hirayama公司;高速冷冻离心机,湖南湘仪离心机仪器有限公司.
1.2 实验方法
1.2.1 ARTP诱变方法
1)菌悬液制备:取1 mL活化菌液于1.5 mL的离心管中,5 000 r/min离心5 min后去除上清液,加入1 mL的无菌生理盐水震动摇匀,制成菌悬液备用.
2)ARTP诱变流程:供电220 V,50 Hz,电压波动范围≤5%,气体流量选用10SLM,功率200 W.ARTP进行诱变处理的处理时间分别为0、30、60、90、120、150、180 s.
3)致死率计算:梯度稀释涂布于MRS固体培养基上,培养48 h后进行菌落计数,按照公式:致死率=(对照组-诱变菌落数)/对照菌落数×100%,计算致死率.
4)筛选:在MRS固体培养基中添加临界浓度的亚油酸(0.5%),将经ARTP诱变后的菌液均匀涂布在平板上,培养一定时间后,挑取平板上生长较好的菌落进行增殖培养并测定酶活[10].
5)遗传稳定性实验:突变株进行7次传代培养,测定每一代菌株的酶活,观察其稳定性.
1.2.2 酶活测定方法
1)粗酶液制备
培养好的菌液4 ℃、6 000 r/min,离心20 min,弃上清液,加入适当的的磷酸盐缓冲液(pH6.0)清洗两次,4 ℃、8 000 r/min,离心10 min,弃上清液,得含酶菌体.将所得含酶菌体加入石英砂研磨后加入5 mL的磷酸盐缓冲液均质10 min,即得菌体重悬液.菌体重悬液加入1%溶菌酶冰水浴中进行间歇破碎15 min(破碎3 s,间歇3 s,超声功率250 W),将破碎菌液于4 ℃、10 000 r/min,离心20 min后,收集上清液,即为粗酶液.
2)共轭亚油酸检测
采用分光光度法以正己烷为参比在233 nm处测定样品的吸收值.以共轭亚油酸质量(X)为横坐标,吸光值(Y)为纵坐标绘制出标准曲线,其回归方程为Y=0.008 2X+0.011 1,R2=0.999 7.根据标准曲线,计算出样品中共轭亚油酸的含量.
3)共轭亚油酸酶活力的测定
在50 mL三角瓶中加入5 mL的粗酶液、100 mg的亚油酸和少许Tween-80,于37 ℃下缓慢振摇1 h,加入10 mL正己烷萃取共轭亚油酸,经4 ℃、10 000 r/min,离心20 min后,取正己烷层,测定其含量.亚油酸异构酶活力单位U定义为:在上述酶活力测定方法下,1 min内生成1 μg CLA 所需的酶量.
酶活力(U/mL)=(K×G)/(V×T),
式中:K为酶液稀释倍数,G为所生成的共轭亚油酸量(μg),V为吸取酶液体积(mL),T为保温时间(min).
2 结果与分析
2.1 碳源及其质量分数对突变株产酶的影响
由图1可知,葡萄糖是最佳碳源,当葡萄糖质量分数为2%时,B5突变株酶活力最高,达到815.4 U/mL.碳源在菌株的生长、繁殖及代谢活动中起着至关重要的作用,在含有充足便于吸收的碳源的环境中,微生物生长繁殖旺盛,代谢水平高,更容易生成丰富的产物.B5突变株利用葡萄糖的效率要显著高于双糖和多糖,说明B5突变株对单糖的利用率要高于多糖.
2.2 氮源及其质量分数对突变株产酶的影响
由图2可知,牛肉膏是最佳氮源,当添加2%的牛肉膏时,B5突变株酶活力最高,达到774.2 U/mL.氮源作为微生物细胞蛋白质的组成部分,对微生物的生长和酶的合成具有重要作用,添加适当质量分数的有机氮源牛肉膏能促进菌体生长和亚油酸异构酶的合成.
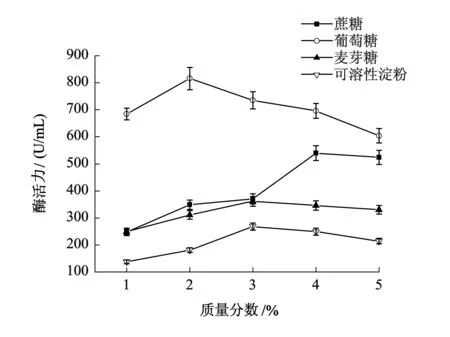
图1 碳源及其质量分数对产酶的影响
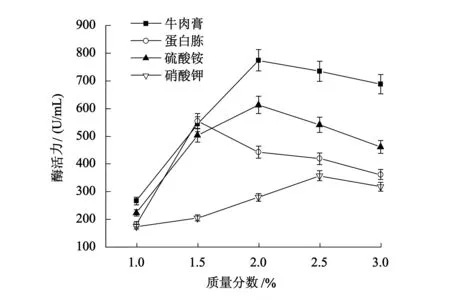
图2 氮源及其质量分数对产酶的影响
2.3 碳氮比对突变株生长和产酶的影响
以葡萄糖和牛肉膏作为碳源和氮源,分别选择碳氮比为1∶0.5,1∶1,1∶1.5,1∶2,1∶2.5.由图3可知,在碳氮比1∶1.5时,B5的生物量和酶活力都达到最高,酶活力为1 083 U/mL.碳源是菌株生长繁殖的保证,氮源是菌体合成蛋白质的来源,合适的碳氮比不仅有利于突变株的生长和代谢,更有利于突变株关键酶的合成[12-13].
2.4 温度对突变株生长和产酶的影响
图4结果表明,温度对突变株产酶的影响显著,当温度低于37 ℃时,随着温度的升高,菌体的生物量和产酶量也在升高.37 ℃时酶活力为1 125 U/mL,生物量为2.344,均达到最大值.继续提高培养温度,B5的生长和代谢都受到抑制,产酶量和生物量均呈快速下降趋势.
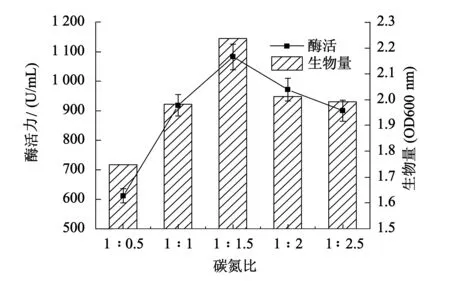
图3 碳氮比对产酶的影响
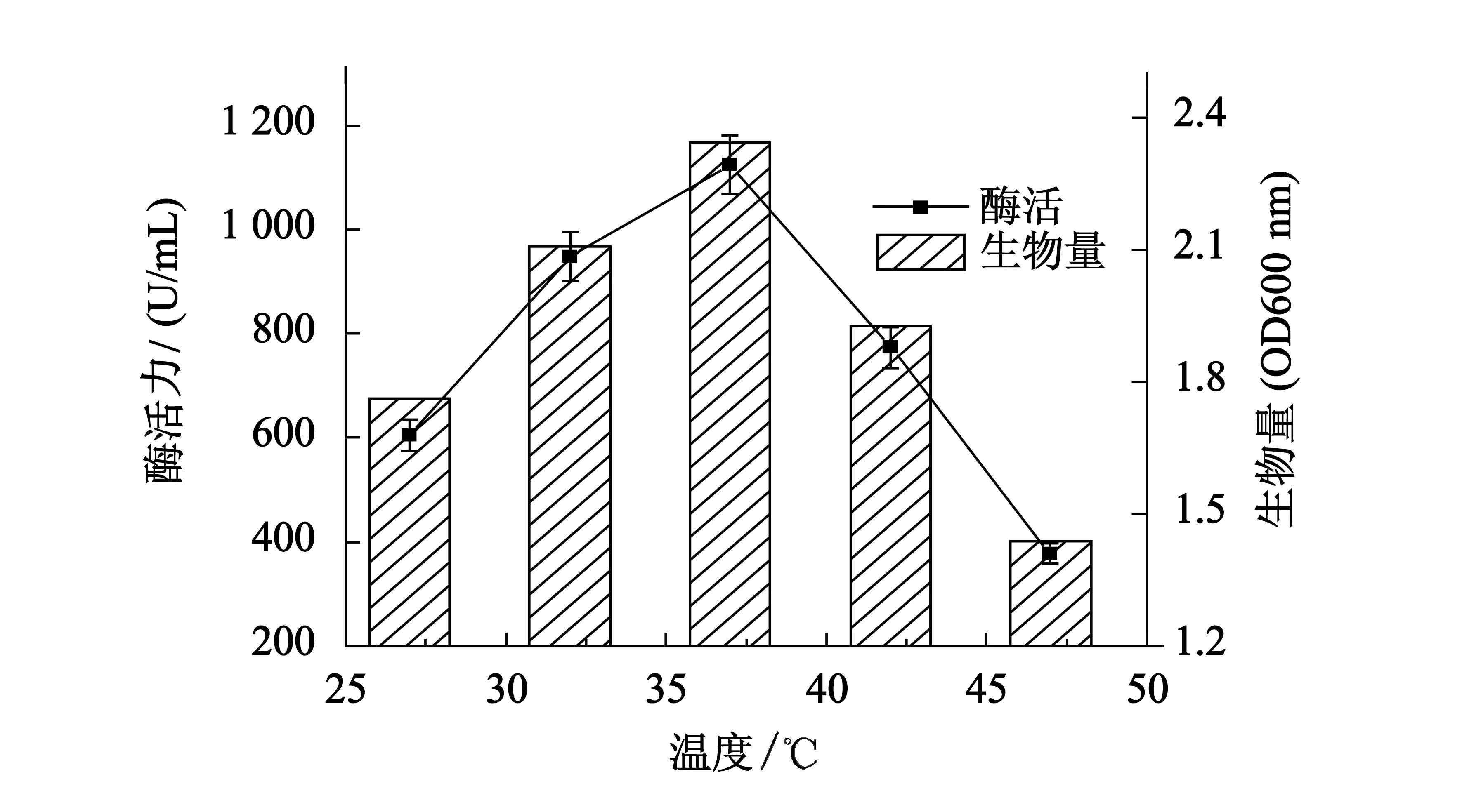
图4 温度对产酶的影响
2.5 培养时间对突变株生长和产酶的影响
B5突变株生长和产酶特性如图5所示,在0~9 h内,菌株产酶量很低,在9~24 h内,菌株开始迅速产酶,在培养36 h时,产酶量达到最大,这是因为亚油酸异构酶是一种诱导酶,当生长环境中含有亚油酸时,需要一定时间诱导突变株中亚油酸异构酶基因的表达,从而开始合成大量的亚油酸异构酶.随着培养时间的进一步延长,产酶量略有下降,这是由于进入静止期,菌体开始自溶,酶合成减少.因此,本实验选择36 h作为B5突变株的最佳培养时间.
2.6 初始pH对突变株生长和产酶的影响
不同初始pH值下B5突变株的产酶及生长情况如图6所示.当初始pH为5.5时,突变株的酶活力和生物量均达到最高,酶活力为1 137.6 U/mL,生物量为2.359;pH值过高或过低均会不同程度抑制菌株的生长和产酶,因此控制合适的初始pH值,对发挥突变株最大的产酶能力非常重要.

图5 突变株的生长和产酶的关系
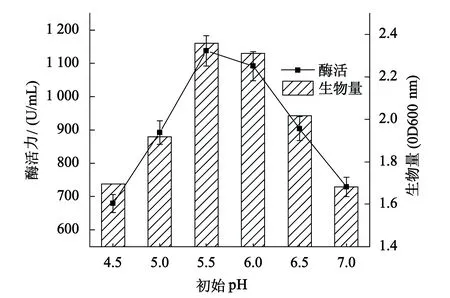
图6 初始pH对产酶的影响
2.7 产酶条件优化与响应面分析
通过对单因素实验结果进行分析,可知培养温度、培养基初始pH、培养基碳氮比是影响B5产酶最显著的3个因素,设定其它条件为:2%葡萄糖,2%牛肉膏,培养时间36 h.设计3因素3水平响应面实验,并通过响应面分析,分析结果见图7~图9.
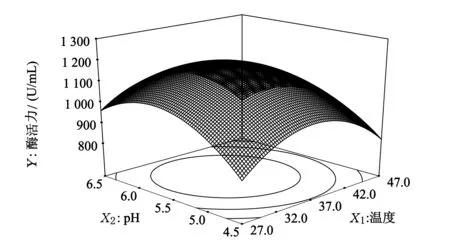
图7 温度和pH的交互作用

图8 碳氮比和pH的交互作用
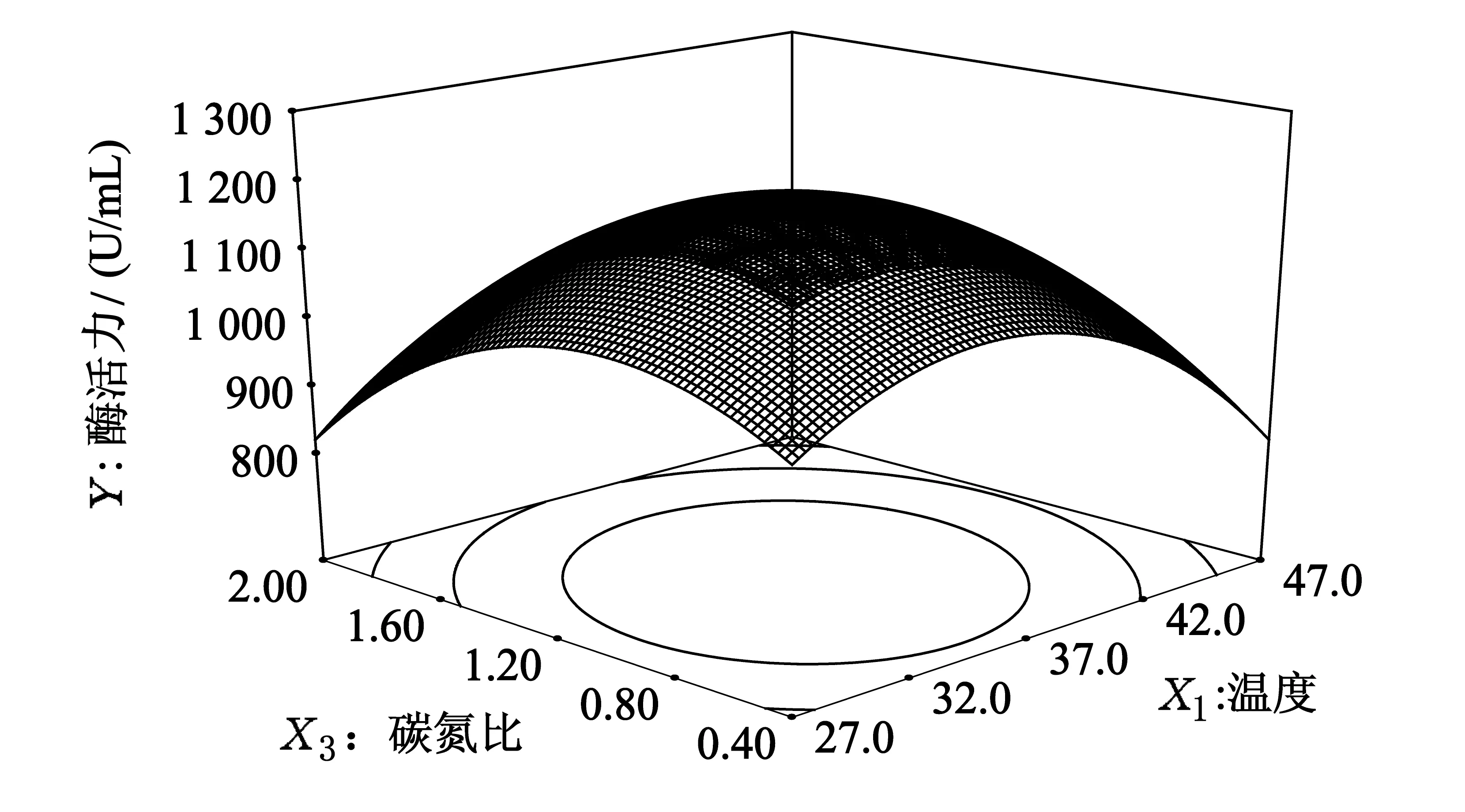
图9 碳氮比和温度的交互作用
由图7可知,设定碳氮比为0时,酶活力随着初始pH的增大先增大后减小.当初始pH较低时,酶活力随着温度的增加先增大后减小.根据酶活力的变化可以看出,初始pH对酶活力的影响大于温度,但两者之间的交互作用不显著.
由图8可知,设定温度为0时,酶活力随着初始pH的增大先增大后减小.当初始pH较低时,酶活力随着碳氮比的增加先呈现上升趋势,后趋于稳定并逐渐开始减小.因此,在一定温度条件下,合适的碳氮比和初始pH有利于突变株产酶量的提高.
固定初始pH在零水平上,酶活力随着温度的增大先增大后减小.当温度较低时,酶活力随着碳氮比的增加先增大,后逐渐开始减小.温度与碳氮比之间存在明显的交互效应(P=0.003 8<0.05),因此,在一定初始pH下,合适的碳氮比和温度有利于突变株产酶量的提高,2个因子在所选择的范围内可以产生最佳响应值.
3 结论
通过对嗜酸乳杆菌进行ARTP诱变处理,得到一株遗传稳定的高产突变株B5,而B5的生长及产酶受多种发酵条件的影响.通过单因素试验结合响应面试验,获得了B5的最佳发酵产酶条件:培养温度36.4 ℃,初始pH5.6,碳氮比1∶1.5,培养时间36 h.亚油酸异构酶产量达到1 214.1 U/mL,是原始菌株的2.84倍.
猜你喜欢
露天采矿技术(2020年6期)2021-01-05 00:37:54
祝您健康(2019年7期)2019-07-12 03:11:52
天然产物研究与开发(2018年9期)2018-10-08 03:25:38
山东工业技术(2016年15期)2016-12-01 05:31:50
西南农业学报(2016年5期)2016-05-17 05:42:39
中国酿造(2016年12期)2016-03-01 03:08:19
应用海洋学学报(2015年3期)2015-11-22 07:39:30
医学研究杂志(2015年8期)2015-06-22 14:00:54
中国食用菌(2013年2期)2013-11-21 03:37:52
食品研究与开发(2012年10期)2012-12-03 05:45:10
