
基于SSR标记的桃种质资源遗传多样性研究
2018-11-16 09:13凌士鹏林贤锐沈建生
江西农业学报 2018年11期
凌士鹏,孙 萍,林贤锐,沈建生
(浙江省金华市农业科学研究院,浙江 金华 321000)
桃是蔷薇科(Rosaceae)李属(Prunus)植物,起源地是中国的西部地区,分布点主要是欧、亚、美、澳、非等五大洲,其种植范围特别广、适应性比较强、种质资源极为丰富[1]。桃经过多年的自然选择和人工驯化种植,形成了许多的类型和品系,这些资源为进一步改良桃的相关性状提供了基础,了解这些资源是利用的前提[2]。目前,相关研究表明:我国拥有北京、南京、郑州3个国家级桃种质资源圃以及很多地方桃种质资源圃,收集并保存了6个种,共2000多份种质资源[3],桃作为世界上栽培最为广泛的落叶果树之一,现已遍布全球,居核果类果树之首[4]。自20世纪80年代以来,国内很多学者对桃亚属种间分类进行了研究,目前已经在形态学[1]、酶学[5]、孢粉学[6]、细胞学[7]、分子标记[8-11]等方面获得了一些桃资源分类及亲缘关系等的结论和观点。但是研究发现,从形态学方面对品种进行分类需要看经验,因此对于一些种间的杂种进行归类十分困难,更无法判断其起源;另外,利用同工酶、染色体核型等试验手段也同样面临着更多的问题。因此开展桃分子标记辅助分类具有重要意义。
简单序列重复(Simple Sequence Repeat,SSR)标记的特点是等位基因变异数多、信息含量及多态性高,稳定性、重复性好,属共显性遗传,特异性强,并且在种属间有良好的通用性[14],是目前果树上鉴定种质资源、分析遗传多样性及亲缘关系应用中最为广泛的分子标记。同时,Verde等[12]基于桃不同器官转录组数据研究后上传至NCBI共80797条ESTs序列,为挑选更好的桃SSR引物提供了丰富的资源。笔者所在课题组已于前期对部分桃种质资源进行了调查、收集保存,因此在此基础上,本研究利用SSR分子标记技术对收集保存的桃种质资源进行遗传多样性的评价,以期为后续桃种质资源的综合利用提供借鉴。
1 材料与方法
1.1 植物材料
于各地收集桃地方品种(品系)共53份,嫁接于浙江省金华国家高科技农业园区的桃李品种园,详细内容见表1。试验材料为健康幼嫩叶片,采集并分别置于采集袋内,同时利用变色硅胶迅速干燥,带回实验室,挑取干燥的叶片进行DNA提取。
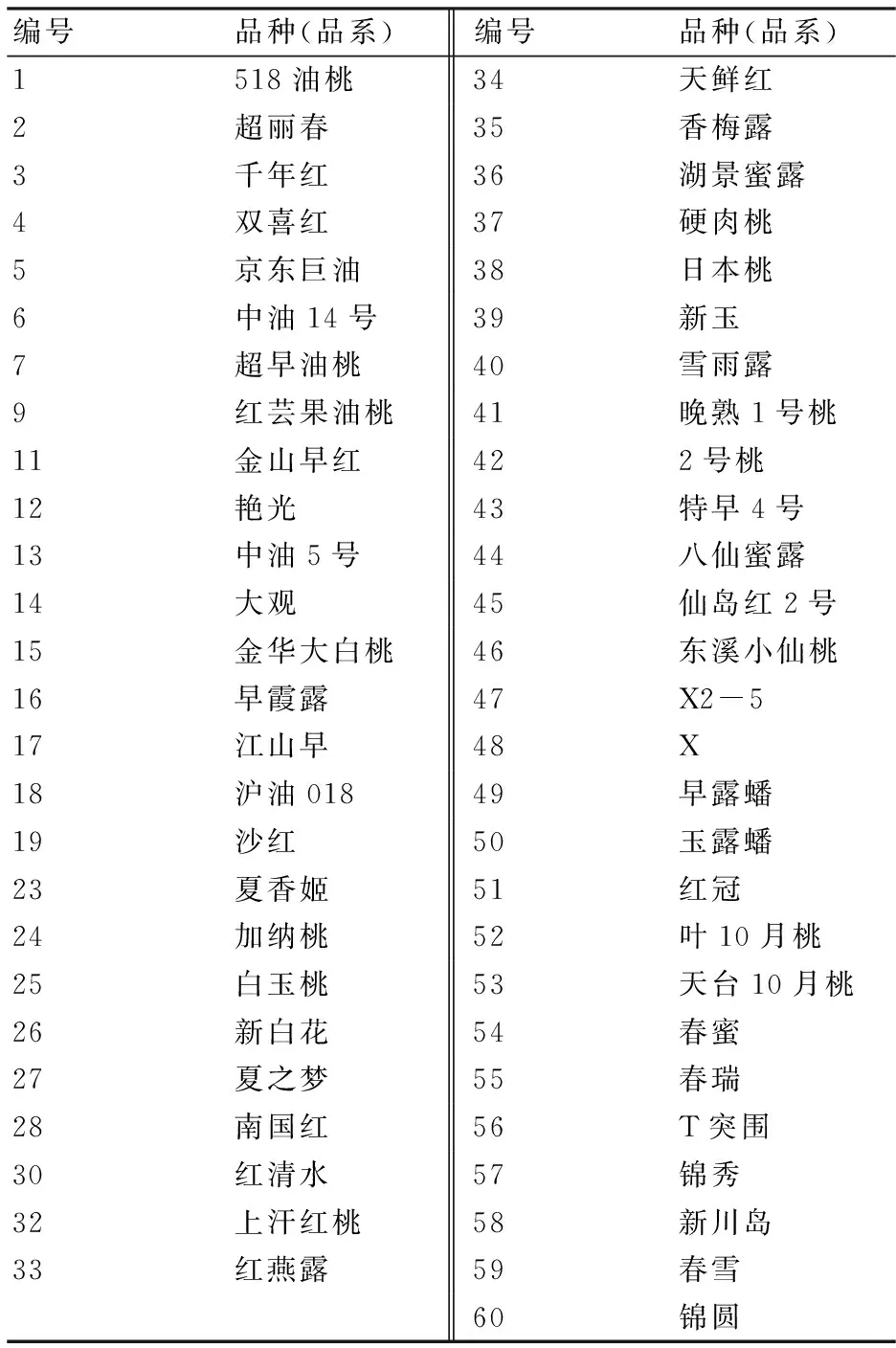
表1 供试的桃品种(品系)
1.2 方法
1.2.1 DNA的提取 采用CTAB法[13]从采集的叶片中提取总DNA,用双蒸水稀释到10~30 ng/μL,储存于-20 ℃冰箱中备用。
1.2.2 SSR标记 随机抽取8个桃样品的DNA模板,选用56对SSR引物进行PCR扩增,将扩增产物利用6%的聚丙烯酰胺凝胶电泳约2.5 h,后银染显色。从中筛选出12对扩增条带清晰、稳定的引物进行SSR扩增,详见表2。试验前对筛选好的12对引物进行荧光修饰,即在单向引物的5′端或3′端加上荧光修饰基团,便可实现微卫星多态性的荧光半自动检测。荧光引物由上海英骏生物技术有限公司合成。
PCR 反应体系及SSR扩增程序详见孙萍等[17]2017年发表的文章。扩增结束后对PCR产物用灭菌的双蒸水稀释100~200倍,利用ABI 3130测序仪进行毛细管电泳检测;数据采集采用Gene Mapper 4.0仪器在电泳结果60~350 bp片段大小范围内进行。
1.3 数据分析
将测序的结果导入Genetic Profile v1.0软件中,参照峰值的强度及分子内标大小,确定微卫星位点扩增片段的区间,对片段大小进行统计,如果出现多峰型的样品,则需重新进行PCR试验后重新检测,以Excel格式保存统计结果,以备后续分析[17]。
利用遗传数据分析软件GenAlEx 6.3[18]计算多态性等位基因数(Na)、SSR位点的有效等位基因数(Ne)、Shannon多样性指数(I)、观察杂合度(Ho)、期望杂合度(He)等遗传多样性指标。
用Population 1.2[19]软件对53个桃种质资源进行Neighbor-Joining聚类分析,并描绘和修改聚类图[20]。
2 结果与分析
2.1 SSR扩增产物的多态性
从表3可以看出,12对SSR引物在所有桃样品中的多态性等位基因数(Na)从6到11不等,等位基因数平均值为7.917。有效等位基因数(Ne)从1.773(CPPCT042)到4.657(BPPCT026),平均值为3.399。其中,引物CPPCT022扩增出的等位基因数为11个,可以区分53个品种中的22个品种;多态性较好的引物为CPPCT022、BPPCT038、CPPCT026、BPPCT025、UDP96-005,能区分几乎所有53个桃地方品种。
在本研究中,观察杂合度(Ho)为0.436(CPPCT042)~0.922(BPPCT038),平均值为0.657。期望杂合度(He)为0.436(CPPCT042)~0.785(BPPCT026),平均期望杂合度为0.678。Shannon多样性指数(I)为0.939(CPPCT042)~1.698(BPPCT038),平均值为1.409。反映了所收集保存的桃种质资源的遗传变异程度较高,遗传多样性较高。

表2 SSR引物及扩增的长度范围

表3 53个栽培桃品种12对引物的遗传参数
2.2 聚类分析
通过SSR数据对53个地方桃品种进行聚类分析,构建亲缘关系图(图1)。从聚类分析图可以得出:53个地方桃品种分成了3组,第一组中包括大观、江山早、加纳桃、红清水等11个品种,其中大观和江山早的相似系数为0.96,晚熟1号桃和2号桃的相似系数为0.98,说明它们之间遗传关系很近;第二组包括了35个品种,其中518油桃和千年红的相似系数为1.00,即为同物异名的现象;第三组包括了其余的9个品种,其中新玉和东溪小仙桃的相似系数为0.94,也表现出了很近的亲缘关系。

图1 基于Nei的遗传距离的Neighbor-Joining聚类分析图
3 小结与讨论
3.1 遗传多样性分析
杂合度的高低及等位基因数的多少是反映所选用SSR位点鉴别植物基因型能力的重要指标[17]。本研究结果表明:所选用的12个SSR引物其多态性较高,可以用于地方桃品种的鉴定和遗传多样性分析,另外也表明了笔者前期所收集保存的地方桃种质资源遗传变异较为丰富。
12对引物在所有样品中等位基因的平均值为7.917,平均有效等位基因数(Ne)为3.399,平均观察杂合度(Ho)为0.657,期望杂合度(He)的平均值为0.678,与史红丽等[21]的研究结果相似(Ne=3.4),比Ghaffari等[22]在扁桃上的研究结果稍低(Na=8.8,Ho=0.7,He=0.79)、但显著高于Aranzana等[23]在桃上的研究结果(Na=6.6~7.3,Ho=0.35,He=0.50~0.55)。另外,本研究中桃种质资源的遗传多样性水平与杏[24](Na=3.5,Ho=0.58)、樱桃[25](Na=3.5~6.0,He=0.49~0.66)等其他李属植物的研究结果相比,有明显差异,表现出较高的遗传多样性水平。这为桃种质资源圃的建立奠定了物质基础。
3.2 遗传结构分析
基于Nei距离的Neighbor-Joining聚类分析清楚检测到桃供试样品的遗传结构:53个地方桃品种分成了3组,其中第一组中的大观和江山早均为早熟桃品种,形态学性状相近;晚熟1号桃和2号桃均为晚熟桃品种,形态学性状也极其相近;另外,第一组中的加纳、红清水、上汗这3个桃品种均为日本选育的品种。第二组的518油桃和千年红的相似系数为1.00,为同物异名的现象;春瑞、春雪、春蜜、突围这4个桃均为早熟桃品种,形态学特征相似;红芸果、艳光、中油5号、超丽春、千年红、京东巨油、超早油桃这几个品种均为油桃品种,聚在一起;红冠、夏香姬、沙红、雪雨露这4个品种形态学特征相似,聚在一起。第三组中新玉和东溪小仙桃的相似系数为0.94,也表现出了很近的遗传关系。
在不同的桃类群中,大部分遗传关系较近的品种以及部分形态学性状相近的品种聚在一起,说明其聚类分析结果与桃系谱以及生物学特征具有一定的关联性,但也不是完全一致的。部分形态学性状相差较大的桃品种,如锦绣和红燕露、天鲜红在聚类图中聚集在一起,说明这3个桃品种之间的基因型存在相互渗透的现象,亲缘关系较近。造成这种现象的原因可能是控制桃肉色性状的基因发生了几个碱基的变化,而通过SSR分析检测不到碱基的微小变异,也可能与桃本身遗传结构相对复杂有关。
本研究结果也显示出聚类结果与桃品种的地理分布之间存在一定相关性,如日本桃品种、部分地方品种分别较为集中的聚在一起。但也有部分来自不同地区甚至地理距离较远的桃品种,其遗传距离表现非常相近,造成这种现象的原因可能与品种所处的环境条件相似,或者人工干预较为相近有一定的关系。因此在选择杂交亲本时,应注重扩大地域选择范围,这对于提高遗传多样性具有明显意义。
另外,研究还存在着普通桃和油桃聚为一类的交叉现象,说明各桃品种之间通过多年的自然选择,以及杂交育种过程中基因之间相互渗透,从而形成了较为丰富的遗传多样性,同时通过生态条件的变化以及人工干预,形成了不同的品种(品系)类型。
本研究采用SSR标记来研究桃种质资源的遗传多样性,结果表明:SSR标记是一种经济、可靠的分子标记技术。从本研究的结论可以看出,所收集到的桃种质资源遗传变异较为丰富,遗传多样性水平较高,对今后桃种质资源的保存和利用具有一定的参考价值。但本研究发现部分性状差别较大的地方桃品种在聚类图上聚为一类,这有待于进一步研究和论证。
猜你喜欢
川北医学院学报(2022年6期)2022-06-24
中国药学药品知识仓库(2022年9期)2022-05-23
昆明医科大学学报(2021年12期)2021-12-30
智慧健康(2021年17期)2021-07-30
种子(2021年3期)2021-04-12
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
遵义医科大学学报(2020年6期)2020-02-05
中国临床医学影像杂志(2019年2期)2019-04-25
校园英语·下旬(2017年7期)2017-07-14
科技视界(2016年27期)2017-03-14
