
H9N2亚型禽流感病毒糖基化位点突变株的构建与生物学特性研究
2018-07-11 05:51:32廖昌韬曾凡桂严专强覃健萍曹永长谢青梅
动物医学进展 2018年7期
廖昌韬,曾凡桂,严专强,覃健萍,3,曹永长,3,谢青梅,陈 峰,,3*
(1.广东温氏食品集团股份有限公司,广东新兴 527439;2.华南农业大学动物科学学院,广东广州 510642;3.广东省畜禽健康养殖与环境控制企业重点实验室,广东新兴 527400)
H9N2亚型禽流感病毒(Avian influenza virus H9N2 subtype,H9N2 AIV)属于低致病性毒株,除了造成禽呼吸道疾病和引起蛋鸡与种鸡的产蛋量下降外[1],还容易与其他病毒或细菌发生共感染,对生产危害极大。随着疫苗的持续使用及养殖环境的不断变化,流感病毒的变异也不断增强,防控难度不断加大[2]。
近年来,对于流感病毒HA位点的变化及其功能改变的研究主要专注于人流感病毒和高致病性AIV。在此之前,针对H1亚型、H3亚型和H5亚型AIV的HA蛋白糖基化位点功能研究表明,缺失或者增加糖基化位点能够影响病毒毒力和免疫逃逸功能[3-4]。而根据现有的报道和数据分析显示,H9N2亚型AIV的HA蛋白第200位N-糖基化位点存在大量的缺失现象,并且具有范围增大的趋势[5]。因此,本研究对本实验室分离纯化的H9N2 AIV流行毒株HA蛋白第200位N-糖基化位点进行突变,利用反向遗传学技术构建了两株重组病毒株并进行初步的探索。
1 材料与方法
1.1 材料
1.1.1病毒株、细胞、质粒及鸡胚Influenza A virus H9N2 A/Chicken/Guangdong/WZP12/2013(KT023025.1),广东省畜禽健康养殖与环境控制企业重点实验室分离鉴定,自然缺失第200位N-糖基化位点,缩写为WZP。大肠埃希菌DH5α感受态细胞与pMD18-T载体,宝生物工程(大连)有限公司产品;双向表达质粒pHW2000质粒、6个内源基因重组表达质粒及293T细胞,中山大学曹永长教授馈赠;MDCK细胞系,本实验室保存;9日龄~11日龄SPF鸡胚,购自新兴大华农禽蛋有限公司。
1.1.2主要试剂Prime Script○ROne Step RT-PCR Kit、pMD18-T载体、DNA Marker DL 2 000、10×Loading Buffer 、EXTaq酶、Premix高保真酶、A-tailing试剂盒,宝生物工程(大连)有限公司产品;核酸染料EB替代物Goldview、琼脂糖,宝泰克生物科技有限公司产品;AxyPrep病毒DNA/RNA提取试剂盒、DNA纯化/回收试剂盒,爱思进生物技术(杭州)有限公司产品;BsmBΙ内切酶、BsaΙ内切酶、T4连接酶、ExTaq酶,NEB(北京)有限公司产品;Endo-Free Plasmid mini Kit Ⅱ转染级质粒提取试剂盒,北京美科美生物技术开发有限公司产品;转染试剂X-tremeGENE9,上海罗氏制药有限公司产品。
1.1.3引物设计及合成参考Hoffmann E等[6]建立的反向遗传系统设计引物,分别在HA基因和NA基因片段添加酶切位点,引物序列为:BmHA-F:5′-CCTTCGTCTCAGGGAGCGAAAGCAGGGG-3′(BsmBⅠ),BmHA-R:5′-GCATCGTCTCGTATTAGTAGAAACAAGGGTGTTTT-3′(BsmBΙ);BaNA-F:5′-CCTTGGTCTCAGGGAGCGAAAGCAGGAGT-3′(BsaΙ),BaNA-R: 5′-GCATGGTCTCGTATTAGTAGAAACAAGGAGTTTTTT-3′(BsaΙ)。根据AIV HA基因序列设计糖基化位点突变引物:HA-200-F:5′-ATAAATAGGACCTTCAAACCA-3′,HA-200-R:5′-TGGTTTGAAGGTCCTATTTAT-3′。
1.2 方法
1.2.1病毒HA基因与NA基因重组表达质粒的构建以WZP毒株为HA与NA基因供体,通过PCR反应克隆目的片段,分别将其插入到pMD18-T载体中进行测序鉴定。以其为模板,分别用定点突变引物BmHA-F/BmHA-R和BaNA-F/BaNA-R进行PCR扩增,将目的片段分别插入到pHW2000载体中,测序之后得到糖基化位点突变的HA基因和NA基因重组表达质粒。
1.2.2重组病毒的拯救将流感病毒8个基因片段的表达质粒按1∶1的比例混合均匀。按照转染试剂盒说明书方法转染293T细胞,培养36 h,每孔加入5 μL TPCK胰酶,培养12 h后冻融3次。加入双抗溶液处理1 h,接种SPF鸡胚,孵育48 h后收集鸡胚尿囊液,检测血凝效价,并通过PCR检测鉴定。
1.2.3HA基因三维结构的预测用Edit Seq软件将核酸序列转化为氨基酸序列,然后将这段氨基酸序列上传到借助SWISS-MODEL预测三维蛋白结构网站(http://swissmodel.expasy.org/interactive),获得HA基因的三维蛋白结构文件。用Pymol软件对获得的三维蛋白结构文件进行可视化操作和修饰,对比氨基酸突变位点在三维蛋白结构中的变化。
1.2.4病毒洗脱试验由于各病毒株NA基因片段相同,因此比较不同AIV病毒株在鸡红细胞上的洗脱时间可以反映出不同HA基因病毒株结合红细胞的能力。将50 μL病毒2倍倍比稀释为相同的HA效价,与等体积5 mL/L的鸡红细胞在96孔血凝板中混合后置于4℃孵育1 h,每个样品设置3个重复。然后将血凝板置于37℃孵育,每隔30 min观察记录一次HA效价,持续记录6 h,分别以37℃孵育时间为横坐标,HA效价为纵坐标绘制曲线图。
1.2.5病毒HA热稳定性试验将试验病毒株的尿囊液稀释为相同的血凝效价,分别在不同温度下(37℃~58℃,设置每1℃为一个梯度)处理1 h,每个温度梯度3个重复。对处理后的病毒通过测定HA血凝效价来判断HA的热稳定性。
1.2.6病毒鸡胚半数感染量、鸡胚平均死亡时间和生长曲线的测定按照试验标准步骤测定鸡胚半数感染量(EID50)和鸡胚平均死亡时间(MDT),分别以100EID50的病毒剂量接种9日龄~11日龄SPF鸡胚,各接种18枚。每间隔12 h收集1次尿囊液,连续收集至72 h;每个时间点分别收集3枚鸡胚尿囊液作为重复,置-80℃保存。测定各病毒株不同时间点收集的尿囊液病毒HA效价,分别用无菌生理盐水进行10倍倍比稀释,接种9日龄~11日龄SPF鸡胚测定EID50。以病毒接种鸡胚后的时间为横坐标,以各时间点所测定的病毒EID50为纵坐标,绘制各病毒接种鸡胚后的生长曲线。
2 结果
2.1 糖基化位点突变重组质粒的构建结果
以WZP株为HA基因和NA基因供体,构建出HA、NA和HA200 3个双向重组表达质粒。其中HA与NA重组质粒含有WZP株HA基因与NA基因的完整序列,HA200重组质粒改变了HA基因第200位潜在N-糖基化位点,经过测序分析,结果如图1所示,与预期结果相符。
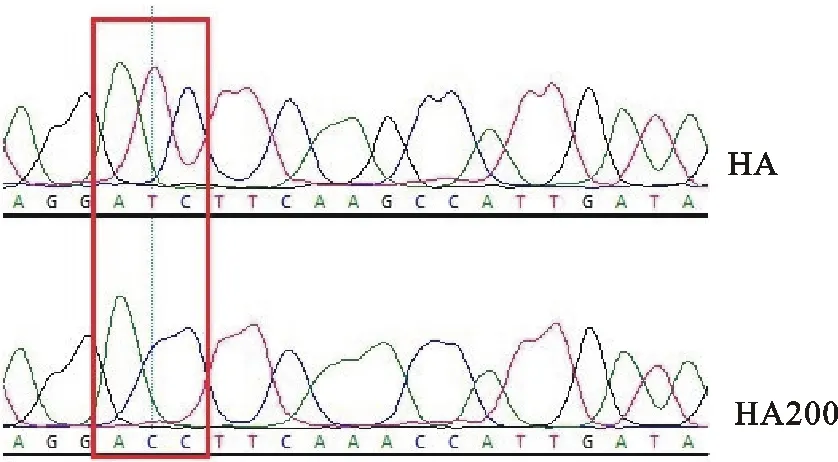
图1 HA基因片段与HA200基因片段序列对比
2.2 病毒拯救及鉴定结果
通过对双表达重组质粒的不同组合,将8种重组质粒共转染293T细胞,拯救出2株重组病毒r-WZP和r-WZP200,其中r-WZP株缺失HA基因第200位N-糖基化位点,r-WZP200株则含有第200位N-糖基化位点。将2株病毒接种SPF鸡胚扩繁3代,每代HA效价(log2)均高于9。通过RT-PCR及测序对病毒8个基因片段进行鉴定,结果表明,其内源基因全部来自病毒株A/Puerto Rico/8/34(H1N1),HA基因与NA基因来自WZP,与预期结果相符。
将重组病毒r-WZP株和r-WZP200株进行大量增殖并浓缩纯化后,通过磷钨酸负染后进行电镜观察。结果表明,在电镜下均能观察到完整的病毒颗粒,并能够观察到囊膜纤突,2株病毒的外观形态无明显区别,表明所救获病毒及相关研究所用材料均为完整的病毒颗粒。
2.3 HA基因三维蛋白结构预测结果
对r-WZP株和r-WZP200株的HA基因三维蛋白结构进行预测,显示HA蛋白第200位糖基化位点位于HA蛋白头部,r-WZP200株因为比r-WZP株多出第200位糖基化位点,其蛋白头部位置可表达出相应的多糖链(图2)。
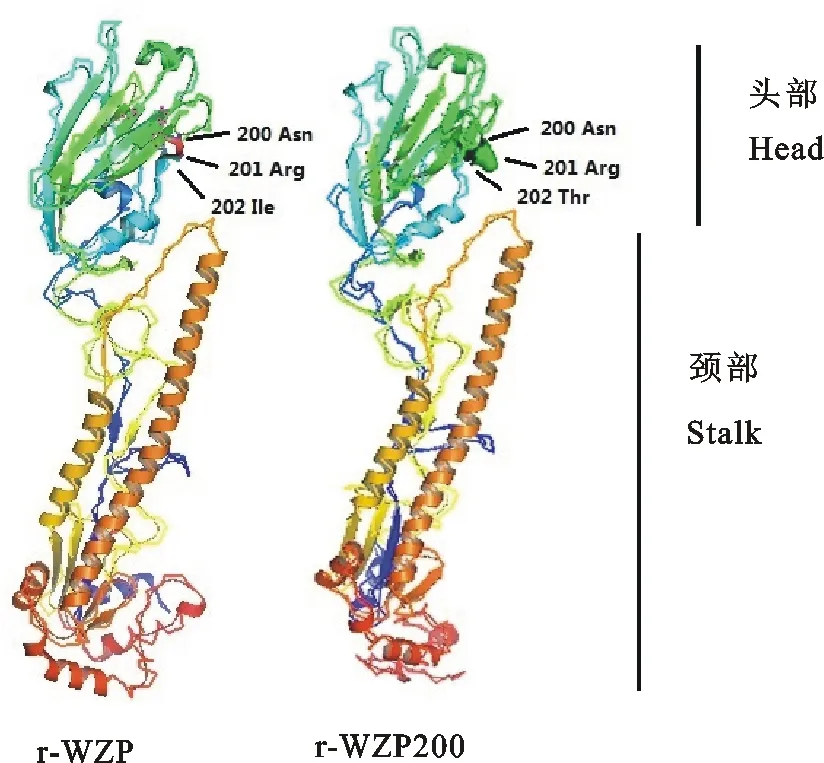
图2 r-WZP株和r-WZP200株的HA基因三维蛋白结构预测
2.4 病毒HA的热稳定性试验结果
2株重组病毒的初始HA效价相同,通过热处理之后HA效价均随着温度的升高而降低,当热处理温度达到56℃时,r-WZP200株HA效价为0,当热处理温度为58℃时,r-WZP株的HA效价变为0。在热处理温度为56℃和57℃时,r-WZP株的HA效价显著高于r-WZP200株(图3)。结果表明,HA基因第200位糖基化位点的缺失能够提高病毒HA的热稳定性。
2.5 病毒洗脱试验结果
测定2株重组毒株在红细胞上的洗脱时间,从而比较二者HA结合鸡红细胞能力的差异。结果显示,r-WZP株和r-WZP200株从鸡红细胞表面解离的时间分别为5.5 h和6 h。在2.5 h~5.5 h期间,r-WZP株的HA效价下降速度很快,而r-WZP200株能够维持更高的HA效价更长的时间,二者差异显著(图4)。表明r-WZP200株具有更强的红细胞结合能力。
2.6 病毒MDT及生长曲线的测定
测定病毒在不同稀释度感染鸡胚后收集的尿囊液HA效价,通过Reed-Muench法计算, r-WZP株和r-WZP200株的EID50/0.1 mL分别为10-8.333和10-7.5。r-WZP株和r-WZP200株的最小致死量为10-4病毒稀释度,根据连续观察记录7 d鸡胚死亡时间,用公式进行计算,r-WZP株的MDT为91.3 h,r-WZP200株的MDT为133 h。
通过测定r-WZP株和r-WZP200株在不同增殖时间点每0.1 mL病毒尿囊液的EID50,绘制出二者在鸡胚尿囊腔中的增殖曲线。结果显示,2株病毒在前期都均呈指数形式增殖,36 h后增殖到平台期,病毒量趋于平稳,并且r-WZP株EID50高于r-WZP200,但差异不显著,表明2株病毒的增殖效率差异不显著(图5)。

图3 不同温度处理对r-WZP株和r-WZP200株HA血凝效价的影响
图4 病毒从吸附的鸡红细胞表面洗脱曲线
3 讨论
HA蛋白是AIV一种重要的表面糖蛋白,而其本身糖基化位点数量和位置的变化会影响蛋白糖链的结构和数量,HA蛋白头部的糖基化位点尤为重要。针对人流感病毒(如H1和H3亚型)的研究发现,糖基化可以帮助病毒逃避宿主的先天性免疫系统,维持病毒的生存[3,7-9]。H5N1亚型AIV HA糖基化位点的缺失能够影响病毒在不同细胞中的增殖,并且影响具有多样性[4,10]。而对于H9N2亚型AIV糖基化位点的研究鲜有报道,主要是通过分子流行病学的调查,监测糖基化位点的变化。本研究针对AIV HA基因第200位N-糖基化位点的缺失进行了功能性的研究。
图5 病毒增殖曲线
通过对AIV HA蛋白三维结构预测分析表明,第200位N-糖基化位点位于HA蛋白头部。研究显示,这一区域的糖基化位点与病毒适应宿主免疫系统的进化过程有关[11],而在病毒与宿主的协同进化过程中,不仅HA蛋白头部糖基化位点的缺失在起作用,共同作用的还有其他一些蛋白的基因位点突变,例如PB2、PB1-F2、PA、M和NS1蛋白的部分位点突变均能够增强病毒毒力和病毒复制效率[9,11]。此外,头部糖基化位点的缺失联合HA蛋白裂解位点或者受体结合位点的变化均能够使病毒更加适应宿主的进化[12]。
流感病毒感染细胞过程中,血凝素(HA)与细胞表面唾液酸受体进行特异性结合,使病毒吸附于细胞表面,进而感染细胞。而神经氨酸酶(NA)具有唾液酸水解活性,能够切断病毒与宿主的结合,起到释放病毒的作用[13]。本研究中r-WZP株和r-WZP200株的NA基因完全相同,细胞吸附洗脱试验结果显示,r-WZP200株具有更强的结合红细胞能力,即第200位N-糖基化位点的缺失显著降低了病毒结合鸡红细胞的能力。该结果证明了第200位N-糖基化位点的变化与病毒结合宿舍受体的能力具有直接关系。
在病毒感染9日龄~11日龄SPF鸡胚的试验中,尿囊腔接种病毒72 h后,收集病毒进行EID50的测定。结果表明,r-WZP株略高于r-WZP200株,但差异不显著。但在MDT的测定中,连续观察7 d的结果显示,2株病毒对鸡胚的最小致死剂量均为10-4稀释度,并且在这个稀释度时,r-WZP株的MDT明显小于r-WZP200株。表明第200位N-糖基化位点的缺失增强了病毒对鸡胚的致死能力。除此之外,同时对分离病毒WZP12株也进行了MDT的测定(本文中未显示数据),发现WZP12株的MDT为83.4 h,小于r-WZP株。
本研究结果表明,HA蛋白第200位N-糖基化位点的缺失降低了病毒结合鸡红细胞的能力,提高了病毒HA效价的热稳定性,增强了病毒对鸡胚的致死性。本研究是首次对H9N2亚型AIV的N-糖基化位点功能进行研究,对H9N2 AIV的致病性研究具有一定的参考价值。
猜你喜欢
中国医学影像学杂志(2023年11期)2023-12-13 09:17:36
心理学探新(2022年1期)2022-06-07 09:16:02
猪业科学(2018年5期)2018-07-17 05:55:18
中国介入影像与治疗学(2017年12期)2017-01-14 18:42:15
山东畜牧兽医(2015年7期)2015-11-23 09:18:30
医学研究杂志(2015年12期)2015-06-10 06:57:46
中国当代医药(2015年8期)2015-03-01 02:01:55
中成药(2014年9期)2014-02-28 22:28:58
中国粮油学报(2014年7期)2014-02-06 01:33:01
现代检验医学杂志(2014年1期)2014-02-06 01:29:26
