
中国壳斗科植物属的空间多样性格局及其指标研究
2018-06-12 09:30:32邱丽氚路丹桂吴晓湲
西北植物学报 2018年4期
邱丽氚,常 虹,路丹桂,吴晓湲,柳 涛
(太原师范学院 地理科学学院,山西晋中030619)
壳斗科植物是构成亚热带常绿阔叶林的主要树种,世界上常绿阔叶林以中国分布面积最大,发育最为典型;中国的落叶阔叶林与世界其他地区不同,栎林是最常见类型[1],可见壳斗科植物对于中国的常绿阔叶林和落叶阔叶林非常重要,将中国壳斗科植物作为研究对象具有重要意义。中国壳斗科植物有7个属,即栗属(Castanea)、锥属(或栲属,Castanopsis)、青冈属(Cyclobalanopsis)、水青冈属(Fagus)、柯属(或石栎属,Lithocarpus)、栎属(Quercus)和三棱栎属(Formanodendron)。
壳斗科植物多样性研究有遗传多样性[2-7]、物种多样性、生态系统多样性[8-10],遗传多样性、生态系统多样性主要采用栎属、栗属、水青冈属等的一些种研究,本研究采用中国壳斗科中所有属种研究物种多样性。
刘茂松等[11-12]对中国壳斗科植物物种多样性的研究是用表格的形式对分布于各省区的属数、种数进行统计,没有采用地理信息系统技术,看不到省区间物种多样性联系即空间多样性。已有研究将地理信息系统技术用于壳斗科植物的研究,但限于一些属、种[13-16],Fang[17]基于中国各县有各种木本植物的分布图,其中包括壳斗科植物种的分布图,没有研究壳斗科植物空间多样性,邱丽氚等[18]研究中国壳斗科植物属的空间分布及多样性,但没有涉及到空间多样性格局指标。
空间格局在植被中研究较多[19-22],但其研究范围较小,没有涉及全国,主要原因是利用遥感影像可获得植被的空间分布,但植被分类比较难,常用植被覆盖度、针阔叶混交林、常绿阔叶林、落叶阔叶林等进行分类,当北方一些森林的建群种明显时,采用建群种分类,如白桦林、樟子松林。植被分类仅涉及较少的物种,对物种的空间格局研究依据物种的分布范围,目前采用植物志、植物标本等处所记载的植物分布地点来确定,最基本的空间分布单元是省或县,省的分布范围比县的大,数据量少,可用列表格的方式研究基于省级的物种多样性。随着地理信息系统技术引入植物空间分布研究,采用县级水平研究物种多样性更容易,不仅可从空间上显示物种分布及空间多样性,而且还可定量研究物种多样性的空间格局,邱丽氚等[23]采用空间多样性格局指标研究中国壳斗科植物空间多样性格局,利用公式计算属、种的空间多样性指标,但没有更详细地研究属内种数的空间多样性格局及属间空间格局的比较。
本研究采用地理信息系统技术,以县为空间数据的基本单元,以壳斗科植物为研究对象,从空间上获取多样性指标,定量地研究中国壳斗科植物的属、类型的空间多样性格局,不仅可获得中国壳斗科植物的物种多样性及其空间分布特点,而且还可获得其空间多样性指标,如不同物种数的面积、斑块数量,探讨空间多样性格局的特点,为空间多样性及其格局研究提供方法、依据,物种多样性的空间分布、面积大小等指标对中国壳斗科植物物种多样性、阔叶林的保护,树木和森林的利用、恢复等森林管理提供依据。
1 数据与方法
1.1 数 据
在中国数字植物标本馆(CVH)网站、中国在线植物志(eFlora)网站、中国树木志、部分省的植物志以及正式发表的论文查找每一种壳斗科植物的具体分布地点,要求每种植物有具体分布的县,将同物异名的种类分布地点合并,分布的点按其所在的县,省区及分布大致范围不能确定其所在的县数据不采用,在种类选择中剔除栽培种类、杂交种类。
1.2 方 法
在ArcGIS9.2软件中制作种的空间数据,包括图形数据和属性数据,图形数据以县为基本单元的中国地图,比例尺为1∶1000000,属性数据为种的分布,然后得到属的种数,制作空间多样性图。

在空间数据中,斑块是由一个县或者是几个相邻的县构成,当斑块的种数相同时,即为同一类型的斑块,如当一个县的种数为1,其周边县的种数不为1,这时斑块是一个县,即一个县形成一个斑块;当几个县的种数均为1,并且这些县相邻,这时斑块是由几个县构成;种数为1的一个县的斑块和几个县的斑块是同一类型的斑块。当种数不同时,形成的斑块则为不同类型,如种数为1的斑块与种数为2的斑块是不同类型的斑块。当斑块种数为1,斑块类型为1,斑块种类为2,斑块类型为2,以此类推。
可看到青冈属前两级的斑块分布区与栗属相似根据空间数据中斑块类型、数目、面积等,获取相关数据,按照空间多样性格局指标公式(表1)计算出属、类型的空间多样性格局指标,分析中国壳斗科植物空间多样性格局,所有计算在Fragstats 3.3软件中完成。在SPSS18软件中计算指标间的相关系数,极显著相关:值>0.9, 显著相关:0.9>值>0.8。
2 结果与分析
2.1 属的空间多样性格局
由表2可知,三棱栎属的PR、NAP均为1,斑块丰富度低,只有1个类型的斑块,只有1个种,连续分布。三棱栎属分布于云南的西南部(图1,a),分布范围明显低于其他属,与表2中它的总面积和总形状指数明显小于其他属相一致。三棱栎属的SHDI、SIDI多样性指数、SHEI、SIEI均度指数均为0,总的最大斑块指数为100,表明没有多样性,与图1,a相吻合。
栗属的PR是2(表2),斑块丰富度低,有2个类型的斑块,其中一个类型有1个种,而另一个类型有2个种,与图1,b图例中的1、2相对应。栗属的总面积、斑块总数、总形状指数均明显高于三棱栎属的相应指标,与图1,b中此属的分布范围大、较为分散、斑块数目多相一致,但它的2个类型的最大斑块指数之和即总的最大斑块指数明显小于三棱栎属,可见属的空间分布范围较大,斑块数目较多,所获得其属的总面积、斑块总数、总形状指数的值均较高,但总的最大斑块指数的值较低。栗属的SHDI、SIDI多样性指数、SHEI、SIEI均度指数均明显高于三棱栎属的相应指标,表明栗属具有多样性,此时斑块丰富度大于1,斑块丰富度影响多样性指数、均度指数。从图1,b中可看到2个类型的分布区均较大, 2个种的斑块周围有1个种的斑块,2个类型的斑块在亚热带地区较均匀分布。
水青冈属的PR为3(表2),斑块丰富度低,有3个类型的斑块,分别是1个种、2个种、3个种的斑块类型,与图1,c图例中的1、2、3相对应。水青冈属的总面积、斑块总数、总的最大斑块指数、总形状指数的值均在三棱栎属与栗属的相应指标值之间,与图1中所看到的内容相一致。水青冈属的SHDI、SIDI多样性指数均与栗属相似,而其SHEI、SIEI均度指数均低于栗属,原因是水青冈属和栗属的斑块丰富度均低,其多样性指数的值相近,水青冈属的前2个类型的斑块分布较均匀,第3个类型的斑块则分布其西北部,造成水青冈属分布不均匀,均度指数低于比栗属。
青冈属的PR是14(表2),斑块丰富度明显高于前3个属,14表示有14个类型的斑块,即1~13的斑块类型分别代表1~13个种,第14个斑块类型有15个种,在图1,d中将斑块类型划分为四级,显示这四级的斑块分布区,以及属的分布区,青冈属分布范围与栗属相似,两属的面积也相近,做如下比较:青冈属的斑块丰富度是栗属7倍,其斑块总数是栗属2倍多,表明斑块类型越多,其斑块总数越多,从图1中可看到青冈属前两级的斑块分布区与栗属相似,但后两级的斑块分布区小,分散分布于青冈属分布区的南部,而每一级中又包括3~4个斑块类型,所以青冈属分布区比栗属更加破碎;青冈属总的最大斑块指数是栗属一半多,总形状指数是栗属3倍多,显示斑块类型越多,其不同斑块类型的最大斑块指数之和缩小,意味着全部或绝大多数不同类型的最大斑块面积缩小,但形状指数之和变大;青冈属SHDI、SIDI多样性指数均明显高于栗属,这与斑块丰富度有关;青冈属SHEI、SIEI均度指数均与栗属相似,分布较均匀,从图1,d中可看到青冈属四级斑块分布图,前两级种数少,面积大,而后两级则相反,前两级决定了青冈属均度指数,综上所述,斑块丰富度可影响斑块总数、总的最大斑块指数、总形状指数、多样性指数。
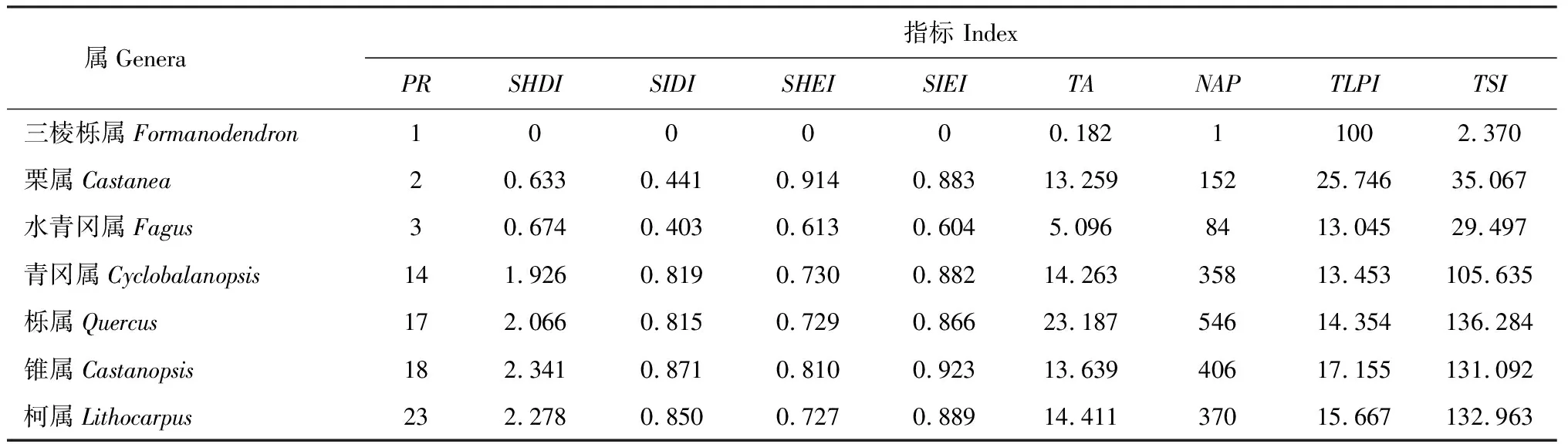
表2 中国壳斗科植物属的空间多样性格局
注:指标及其计算公式和含义详见表1
Note: The formula and meaning of index is detailed in Table 1

图1 中国壳斗科植物中属的空间多样性分布Fig.1 Spatial diversity distribution of every genus in Fagaceae of China
栎属的PR为17 (表2),斑块丰富度略高于青冈属,17表示有17个类型的斑块,即1~16的斑块类型分别代表1~16个种,第17个斑块类型有19个种,在图1,e中显示栎属的分布范围明显大于其它属的,从表2中属的总面积可看到这个特点,同时斑块总数明显高于其他属,其总的最大斑块指数、总形状指数均与青冈属相应指标接近。栎属的SHDI、SIDI多样性指数、SHEI、SIEI均度指数均与青冈属接近。
锥属的PR为18 (表2),斑块丰富度与栎属相近,比青冈属多, 18表示有18个类型的斑块,即1~17的斑块类型分别代表1~17个种,第18个斑块类型有20个种,在图1,f中显示锥属分布范围与青冈属相似,它们的总面积相近,但锥属的分布区更朝南,更加紧密,锥属的其他指标比青冈属的相应指标都高。
柯属的PR是23(表2),斑块丰富度明显高于其他属,23表示有23个类型的斑块,即1~22的斑块类型分别代表1~22个种,第23个斑块类型有35个种,在图1,g中显示柯属的分布范围与锥属的相似,其总面积、总形状指数与锥属相近,但其他指标均略低于锥属的相应指标。
对7个属(表2)做相关性分析时,三棱栎属与其他属不相关,其他属间均呈极显著正相关(P<0.01),其中青冈属、栎属、锥属、柯属这4个属间的相关系数最高,栗属与水青冈属的相关系数较高,与上述结果相一致。
空间多样性格局指标做相关性分析,PR、SHDI、SIDI、NAP、TSI相互之间、NAP与TA之间呈极显著正相关(P<0.01),TLPI与SHEI、SIEI之间呈极显著负相关(P<0.01)。SHDI、SIEI、TA相互之间,TA与SIDI、TSI之间呈显著正相关(P<0.05)。SIDI与TLPI之间呈显著负相关(P<0.05)。斑块类型数目越多,属的空间分布更加多样性,斑块总数越多,总形状指数越高,原因是1个斑块类型有1个或多个斑块,多个斑块类型就有更多的斑块,斑块总数增加,也增加了空间多样性,多个斑块类型或斑块的周长比其任何一个的周长要长,使得总形状指数增加。斑块总数越多,属空间分布的总面积越大,总面积又影响属的空间多样性格局。总的最大斑块指数越大,属的空间分布的均匀度越小,原因是最大斑块越多,优势现象越明显,空间分布更不均匀。
2.2 类型的空间多样性格局
由表2、表3可知,三棱栎属的类型指标(表3)与属的对应指标(表2)的数值相同,如斑块类型与PR的值均为1,APT与TA的值均为0.182,原因是三棱栎属只有1个斑块,这个斑块的各项指标既代表属也代表种,属与种的空间多样性格局相同。
栗属斑块类型分别为1、2,当斑块类型为1时,其所占面积、占属面积的比例、斑块数目、最大斑块指数、形状指数均明显高于斑块类型为2的相应指标(表3),表明斑块类型为1占优势,与图1,b中斑块类型为1分布在斑块类型为2的周围相一致,两个斑块类型呈极显著正相关(P<0.01)。
水青冈属斑块类型分别为1、2、3,每个斑块类型都有所占面积、占属面积的比例、斑块数目、最大斑块指数、形状指数,随着斑块类型从1增加到3,其相应指标依次降低,其间差别较大(表3),与图1,c相一致,斑块类型1、2之间呈极显著正相关(P<0.05),斑块类型3则与其他斑块类型不相关,与图1,c相一致,斑块类型3分布在局部、范围小,与其他斑块类型分布不同。水青冈属的斑块类型所占面积、斑块数目、最大斑块指数、形状指数比栗属相应斑块类型的对应指标均减少,显示水青冈属比栗属相应斑块类型的分布范围缩小,最大斑块也缩小,斑块数目、破碎度均降低,但当斑块类型为1时水青冈属所占属的面积比例高于栗属,优势现象更加明显(表3、图1)。

表3 中国壳斗科植物中3个属的类型空间多样性格局
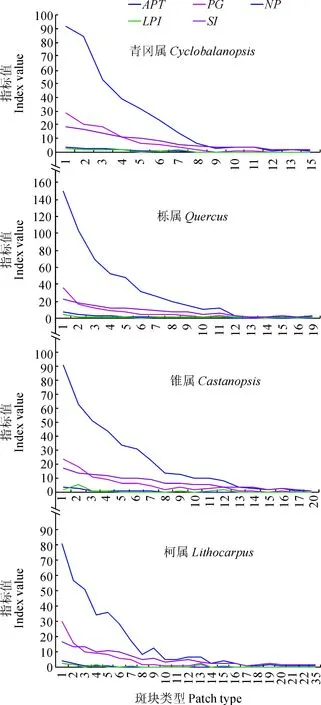
图2 中国壳斗科植物中4个属的类型空间多样性格局Fig.2 Spatial diversity patterns of types in four genera of Fagaceae in China
青冈属、栎属、锥属、柯属的所占面积、占属面积的比例、斑块数目、最大斑块指数、形状指数各指标基本上均随所在属的斑块类型值的增加而相应指标依次降低(图2),与栗属、水青冈属有同样的结果,斑块类型值的增加,意味着种类数量的增加,种类多样性越来越明显,但其分布范围越来越小(图1),使其属所占面积等指标下降,景观优势和破碎度降低。青冈属、栎属、锥属、柯属的斑块类型、类型面积、类型占属面积的比例、斑块数目、最大斑块指数、形状指数做相关性分析,结果显示斑块类型与其他指标呈极显著负相关(P<0.01),说明斑块类型越多,其各项指标均降低,与上述结果相一致。
将青冈属的所有斑块类型进行相关性分析,1、2、3、4、5、6、7(即1~7)的斑块类型相互之间呈极显著相关或显著相关(P<0.01或0.05),9、10、11、12、13、15(即9~15)的斑块类型相互之间呈极显著相关(P<0.01),而斑块类型8只与斑块类型9、10、11、13之间呈极显著相关或显著相关(P<0.01或0.05),可见种数较少(1~7)的斑块类型空间多样性格局相似,只是分布范围缩小,随着种数的增加,在前一个斑块类型的基础上,更向其中心缩小分布区,破碎度降低,种数较多(9~15)的斑块类型也有相似的特点,种数较少与种数较多的斑块类型空间多样性格局差异明显,界线清楚,以斑块类型7、8之间为界,从图1~2中可看到此特点。栎属、锥属、柯属也有类似的特点,只是随着斑块类型的增加,种数较少与种数较多的斑块类型的界线逐渐不明显。4个属种数较少的斑块类型所占面积、占属面积的比例、斑块数目、最大斑块指数、形状指数各指标的值均较大,种数为1~7的斑块类型所占面积之和、斑块数目之和、最大斑块指数之和、形状指数之和分别占其属对应总指标的值均高于57%,其中前两项指标的值均高于80%,可见种数较少的斑块类型构成了属的主要分布区(图1),但后两项指标的值在青冈属、栎属、锥属、柯属中对应指标依次降低,与斑块类型的数量有关。种数较多的斑块类型,其各指标的值均较低,分布范围很狭窄(图1),但物种多样性明显。
3 讨 论
物种多样性是指一个地区内物种的多样性,或从物种组成方面研究群落的组织水平[24],本研究与以往的物种多样性研究有所不同,研究范围很大,覆盖整个中国,其内部细分到县,多样性类型是基于种的数量划分,采用地理信息系统技术不仅从空间上直观地看到物种多样性,而且从分布图上获取斑块数据、计算指标,掌握中国壳斗科植物每一个属的空间多样性、属及其下种数的面积、斑块数目等,定量地分析空间多样性格局及指标间关系,使得物种多样性研究如同植被景观一样进行大范围研究,探讨其空间分布特点,这是本研究的特色。
属、类型的空间多样性格局指标能反映他们各自的空间分布特点,属的斑块丰富度与斑块总数、总形状指数间呈现正相关,类型则是属内的斑块类型,斑块类型与斑块数目、形状指数呈现负相关,前者是属间比较,后者是属内比较。中国壳斗科植物一个属是一个图层,图层中图形数据按其斑块类型划分,不同属图形数据有不同程度的重叠[18],除三棱栎属外,其他属在中国广大的亚热带地区重叠度大,属间的相关系数高,亚热带地区是这些属的主要分布区,绝大多数种类生长于亚热带地区,与壳斗科植物是构成亚热带常绿阔叶林的主要树种相一致[1]。在属的主要分布区内许多种分布范围狭窄,它们局限分布于一个县或几个县,而这些县多为山区,随着海拔高度的升高,温度会降低,为狭域性壳斗科植物提供不同的环境条件,使得壳斗科植物种类多,种数也多,斑块类型高,但其斑块数目较少,形状指数较低。随着种数较多的区域向外扩展,种数逐渐减少,由狭域性种类向广域性种类过渡,广域性种类的分布范围较大,种类少,斑块类型低,但斑块数目较多、形状指数较大。属内斑块类型的空间分布不重叠,种类多的斑块类型被种类少的斑块类型的分布区所包围,具有从多样性中心向外扩展的分布规律,多样性格局指标也能反映此规律。
在青冈属、栎属、锥属、柯属中每一个属种数为1~7的斑块类型所占面积之和、斑块数目之和分别占其属对应总指标的值均高于80%,种数较少的斑块类型构成了属的主要分布区,将这些种类可作为植被恢复、开发利用的壳斗科植物。种数较多的斑块类型,其各指标的值均较低,分布范围很狭窄,但物种呈现多样性,将种数很多的区域作为保护区,以保护壳斗科植物物种资源。
从遥感影像或植被图可获取植被数据,用景观格局指标能分析植被空间格局。当植被建群种明显时,可获得主要层优势种的空间格局,但物种只限于优势种,种类太少,当植被的优势现象不明显时,空间格局不涉及物种。而物种对植物群落、植被的构成非常重要,物种的调查需要巨大的人力、物力,中国已出版许多植物志,其内有植物的分布地点,为物种空间分布研究奠定了基础。物种多样性空间格局的研究不仅能看到物种的空间分布,还能获得数量指标,对掌握中国物种多样性的空间性、定量性非常重要,为物种、植被的保护、利用、恢复,中国自然环境复杂性,高分辨率遥感影像判读植物信息等提供依据。
参考文献:
[1] 武吉华, 张 绅, 江 源,等. 植物地理学(第四版)[M], 北京: 高等教育出版社, 2004.
[2] 李俊清. 中国水青冈种内种间遗传多样性的初步研究[J]. 生物多样性, 1996,4(2): 63-68.
LI J Q. Studies on intra-and inter-species gene diversity of Chinese beeches[J].ChineseBiodiversity, 1996,4(2): 63-68.
[3] KUMAR A, ROGSTAD SH. A hierarchical analysis of minisatellite DNA diversity in Gambel oak (QuercusgambeliiNutt.; Fagaceae)[J].MolecularEcology, 1998,7(7): 859-869.
[4] 郎 萍, 黄宏文. 栗属中国特有种居群的遗传多样性及地域差异[J]. 植物学报, 1999,41(6): 651-657.
LANG P, HUANG H W. Genetic diversity and geographic variation in natural populations of the endemicCastaneaspecies in China[J].ActaBotanicaSinica, 1999,41(6): 651-657.
[5] FERNANDEZ-M. J F, SORK V L. Genetic variation in fragmented forest stands of the andean oakQuercushumboldtiiBonpl. (Fagaceae)[J].Biotropica, 2007,39(1): 72-78.
[6] ROGSTAD S H, PELIKAN S. Restoring the American chestnut: optimizing founder spacing to promote population growth and genetic diversity retention[J].RestorationEcology, 2014,22(5): 668-675.
[7] YING L X, ZHANG T T, CHIU C A,etal. The phylogeography ofFagushayatae(Fagaceae): genetic isolation among populations[J].EcologyandEvolution, 2016,6(9): 2 805-2 816.
[8] ITOW S. Species turnover and diversity patterns along an evergreen broad-leaved forest coenocline[J].JournalofVegetationScience, 1991,2(4): 477-484.
[9] 熊莉军, 郭 柯, 赵常明, 等. 四川大巴山米心水青冈(Fagusengleriana)群落的物种多样性特征[J]. 生态学报, 2008,28(8): 4 007-4 014.
XIONG L J, GUO K, ZHAO C M,etal. Species diversity ofFagusenglerianacommunity in Daba Mountains, Sichuan, China[J].ActaEcologicaSinica, 2008,28(8): 4 007-4 014.
[10] WURST S, KAISER N, NITZSCHE S,etal. Tree diversity modifies distance-dependent effects on seedling emergence but not plant-soil feedbacks of temperate trees[J].Ecology, 2015,96(6): 1 529-1 539.
[11] 刘茂松, 洪必恭. 中国壳斗科的地理分布及其与气候条件的关系[J]. 植物生态学报, 1998,22(1): 41-50.
LIU M S, HONG B G. The distribution of Fagaceae in China and its relationship with climatic and geographic characters[J].ActaPhytoecologicaSinica, 1998,22(1): 41-50.
[12] 刘茂松, 洪必恭. 中国壳斗科的分布格局类型分析[J]. 南京林业大学学报, 1999,23(5): 18-22.
LIU M S, HONG B G. The analysis of distribution pattern of Fagaceae in China[J].JournalofNanjingForestryUniversity, 1999,23(5): 18-22.
[13] 方精云, 郭庆华, 刘国华. 我国水青冈属植物的地理分布格局及其与地形的关系[J]. 植物学报, 1999,41(7):766-774.
FANG J Y, GUO Q H, LIU G H. Distribution patterns of Chinese beech (FagusL.) species in relation to topography[J].ActaBotanicaSinica, 1999,41(7):766-774.
[14] 刘国华, 方精云. 我国栗属物种(Castaneamillissima)地理分布及其空间特征分析[J]. 生态学报, 2001,21(1): 164-170.
LIU G H, FANG J Y. Spatial patterns of chestnut (Castaneamillissima) and its species geographical distribution in China[J].ActaEcologicaSinica, 2001,21(1): 164-170.
[15] 龚 珑, 阎恩荣, 王希华. 常绿阔叶林壳斗科优势种植物地理特征数量分析[J]. 华东师范大学学报(自然科学版), 2008, (6): 63-74.
GONG L, YAN E R, WANG X H. Quantitative analysis on the phytogeographic characteristics of Fagaceae dominant species in evergreen broad-leaved forest[J].JournalofEastChinaNormalUniversity(Natural Science), 2008, (6): 63-74.
[16] XU X T, WANG Z H, RAHBEK C,etal. Geographical variation in the importance of water and energy for oak diversity[J].JournalofBiogeography, 2016,43(2): 279-288.
[17] FANG J Y, WANG Z H, TANG Z Y. Atlas of woody plants in China vol. 1[M]. Beijing: Higher Education Press, 2011.
[18] 邱丽氚, 吴晓湲, 柳 涛.中国壳斗科植物属的空间多样性及差异[J]. 西北植物学报, 2016,36(10): 2 103-2 108.
QIU L C, WU X Y, LIU T. Spatial diversities and differences of all genera in Fagaceae of China[J].ActaBot.Boreal. -Occident.Sin., 2016,36(10): 2 103-2 108.
[19] 邱丽氚, 王尚义. 山西植被空间格局及演替[J]. 植物研究, 2014,34(1): 6-13.
QIU L C, WANG S Y. Spatial patterns and successions of vegetation in Shanxi[J].BulletinofBotanicalResearch, 2014,34(1): 6-13.
[20] 董灵波, 刘兆刚, 李凤日. 大兴安岭盘古林场森林景观的空间分布格局及其关联性[J]. 林业科学, 2015,51(7): 28-36.
DONG L B, LIU Z G, LI F R. Spatial point patterns and associations of forest landscapes in Pangu forest farm in Daxing’an Mountains[J].ScientiaSilvaeSinicae, 2015,51(7): 28-36.
[21] 胡美娟, 侯国林, 周年兴, 等. 庐山森林景观空间分布格局及多尺度特征[J]. 生态学报, 2015,35(16): 5 294-5 305.
HU M J, HOU G L, ZHOU N X,etal. Spatial distribution patterns and multi-scale features of the Lushan forest landscape[J].ActaEcologicaSinica, 2015,35(16): 5 294-5 305.
[22] 徐凯健, 曾宏达, 任 婕, 等. 亚热带典型红壤侵蚀区人类活动对植被覆盖度及景观格局的影响[J]. 生态学报, 2016,36(21): 6 960-6 968.
XU K J, ZENG H D, REN J,etal. Spatial and temporal variations in vegetation cover in an eroded region of substropical red soil and its relationship with the impact of human activity[J].ActaEcologicaSinica, 2016,36(21): 6 960-6 968.
[23] 邱丽氚, 柳 涛, 常 虹, 等. 中国壳斗科植物空间多样性格局研究[J]. 植物科学学报, 2017,35(2): 234-242.
QIU LC, LIU T, CHANG H,etal. Study on the spatial diversity patterns of Fagaceae in China[J].PlantScienceJournal, 2017,35(2): 234-242.
[24] 钱迎倩, 马克平. 生物多样性研究的理论与方法[M]. 北京: 中国科学技术出版社, 1994.
猜你喜欢
英语世界(2023年10期)2023-11-17 09:18:18
自我保健(2021年2期)2021-11-30 10:12:31
妇女之友(2021年9期)2021-09-26 14:29:36
昆明医科大学学报(2020年11期)2020-12-28 00:47:08
科学大众(中学)(2019年3期)2019-05-17 10:04:30
百姓生活(2019年2期)2019-03-20 06:06:16
数理化解题研究(2018年34期)2018-12-27 08:30:04
汽车观察(2018年10期)2018-11-06 07:05:26
数理化解题研究(2018年16期)2018-07-12 02:46:56
少儿科学周刊·少年版(2015年1期)2015-07-07 17:15:12
