
柴达木盆地荒漠灌丛群落谱系结构研究
2018-06-12 09:28:52肖元明杨路存聂秀青李长斌赵晓辉周国英
西北植物学报 2018年4期
肖元明,杨路存,聂秀青,李长斌,熊 丰,赵晓辉,周国英*
(1 中国科学院 西北高原生物研究所,西宁 810008;2 中国科学院 藏药研究重点实验室,西宁 810008;3 青海省青藏高原特色生物资源研究重点实验室,西宁 810008;4 中国科学院大学,北京 100049)
近年来全球变暖、降水格局改变、生境破碎化等因素使生物多样性丧失的形势日益严峻[1-2],生物多样性丧失对人类社会已产生重要影响[3],因此如何有效地保护生物多样性已成为生态学研究的热点问题[4-5]。特别是在生物多样性较低的干旱生态系统,生态系统本身的脆弱性与全球化环境问题双重影响使这些地区的生物多样性保护工作变得更加紧迫[6-7]。理解群落构建过程即群落生物多样性维持机制,是生物多样性保护的重要理论前提[8]。随着最近几十年群落生态学的快速发展,生境过滤、种间竞争、扩散限制等生态学概念相继被提出[9-10],试图阐明群落构建过程及生物多样性分布规律,但这些群落生态学研究大多以物种名来表示群落物种组成,而物种名仅是某一物种的表达手段,其本身不会传递任何有关物种进化或者生态的相似性或者差异[11-12]。例如,群落中物种发生替换,这些物种与原来被替换的物种在功能上可能相似也可能不同或者完全没有关系。因此在全球变化的背景下,迫切需要将群落谱系信息整合进群落生态学研究,从群落中物种之间谱系关系上对群落动态进行分析[13]。
自Webb[11]首次将谱系的方法应用于群落生态学研究并提出度量群落谱系结构的指数(NRI和NTI)[14]之后,国内外众多学者相继开展了许多相关研究来探索群落构建机制,极大地丰富了群落谱系生态学理论。如Swenson等[12]在研究群落谱系时发现群落谱系研究结果与空间尺度有密切的联系。Letcher等[15]对不同演替阶段的群落研究发现随着演替的进行主导群落构建的主要生态过程会发生变化。姜晓燕等[16]利用NRI指数研究了山西霍山植物群落的谱系结构,证明了生态位理论在霍山植物群落构建中具有更加重要的作用。Huang等[17]研究发现:非生物环境因子过滤是中国亚热带森林群落构建的主导生态过程,证明了环境因子对群落谱系有重要的影响。群落谱系的研究方法也为有效的生物多样性保护策略制定提供了新思路。传统的生物多样性保护策略多是基于物种水平的生态学指标,如物种丰富度等[18],而忽略了不同物种之间在进化历史上的差异[19]。研究发现在生物多样性保护策略制定中考虑群落谱系信息能极大地弥补传统保护策略的不足[20],且不受物种分类地位的影响,能更好地反映生态系统[21-22],所以许多学者和生物多样性保护组织呼吁在未来的生物多样性保护行动中要将群落谱系多样性的内容纳入其中[4,20,23-24]。如Devictor等[25]将谱系多样性(Faith’s PD)[26]用于确定某区域优先保护的群落,认为群落中物种之间亲缘关系较远的群落灭绝将会损失更多进化上的多样性。慈秀芹等[23]也认为只有最大化地保存物种进化上的多样性,才能为将来面对全球气候变化提供更多生物多样性演化的潜力。
在以往的谱系群落生态学研究中,大多数学者将研究的焦点放在热带、亚热带森林等物种丰富、气候条件优越的群落或者是中长期的群落演替序列上[11-13,15-17],很少有学者关注柴达木盆地等高海拔、干旱生态系统的群落谱系结构。柴达木盆地位于青藏高原西北部,是中国海拔最高的内陆盆地,对气候变化十分敏感。柴达木盆地稀疏而单一的荒漠植被对于柴达木地区生态环境保护及经济健康发展至关重要[27]。柴达木地区也是世界上极端干旱的地区之一,同时伴有一些河谷(河漫滩)地带,这种地理条件为全球变化背景下研究高海拔干旱生态系统中植物群落谱系结构与水环境因子之间的关系提供了一个理想的天然实验场所。本试验以柴达木盆地荒漠灌丛群落为研究对象,拟探讨柴达木盆地荒漠灌丛群落谱系结构及其与环境因子的关系,旨在为加深对荒漠生态系统群落谱系结构的理解提供科学案例以及为将来相关模型的开发提供更加多元的数据;阐述柴达木盆地灌丛群落物种之间的亲缘关系和群落生物多样性维持机制,为当地生物多样性保护实践提供一定的理论基础。
1 材料和方法
1.1 研究区域概况
柴达木盆地是中国地势最高的内陆盆地,位于青海省西北部(35°0′~39°20′N、90°16′~99°16′E),海拔2 650~6 615 m,面积约2.75×105km2。属大陆性气候,以干旱严寒为主要气候特征。据多年气象资料显示,盆地内多年平均气温3.5 ℃左右,最冷月份1月平均气温-12 ℃,7月气温最高,平均气温16 ℃。降水稀少,多年平均降水量约80 mm。日照时间长,蒸发强烈,多年平均蒸发量约1 500 mm[28]。盆地内地貌类型多样,从盆地边缘至中心分布有高山、戈壁、平原、盐沼、湖泊等地貌类型。盆地主要土壤类型是盐化荒漠土和石膏荒漠土[29]。植被属于典型的荒漠半荒漠植被,结构简单,物种组成单一,以具高度抗旱能力的灌木、半灌木、草本植物为主,分布于砾石戈壁、洪积平原沙漠带[27]。灌木以梭梭(Haloxylonammodendron)、唐古特白刺(Nitrariatangutorum)、多花柽柳(Tamarixhohenackeri)等为主,半灌木以五柱红砂(Reaumuriakaschgarica)、驼绒藜(Ceratoideslatens)、盐爪爪(Kalidiumfoliatum)等为主,草本层以赖草(Leymussecalinus)、芦苇(Phragmitesaustralis)、青海猪毛菜(Salsolachinghaiensis)、垫状凤毛菊(Saussureapulvinata)、青藏狗娃花(Heteropappusbowerii)等为主。
1.2 取样方法
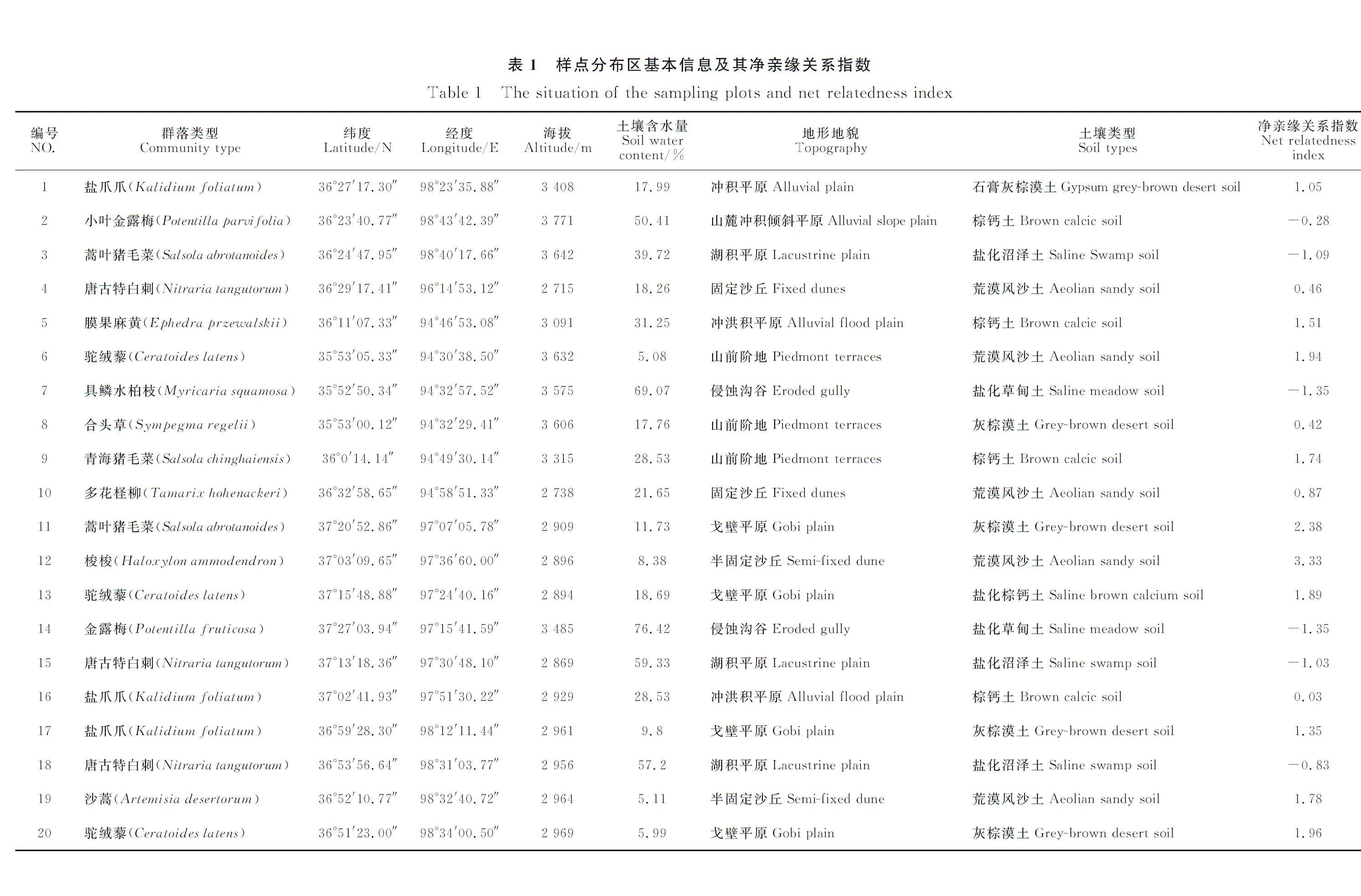
依据《生态系统固碳观测与调查技术规范》[30]进行样地布设。在地势较为平坦、群落类型特征明显、人为干扰较少的条件下,从柴达木盆地东部向西选择了20个具有较强代表性的地段作为样地。样地选定之后,使用手持式GPS记录每个样地的经纬度、海拔,并记录群落类型、生境类型等信息,使用土壤水分仪(TDR)测定土壤水分,在测定时人工挖一个深1 m的土壤剖面,使用TDR分5层测定土壤水分,计算其均值来代表该样地土壤水分状况。随后分别在每个样地有代表性的地段设置3个10 m×10 m样方,样方之间距离保持在5 m至50 m之间。在10 m×10 m的样方内采集灌丛及草本植物标本,每个样地内的3个样方调查的物种名录合并在一起来表示该样地的物种组成。20个样地共计90个物种(包含3种裸子植物),属于23科70属。
1.3 谱系树的构建
物种定名参考《中国在线植物志》(http://frps.eflora.cn/),根据TPL(the Plant List)网站对定名进行规范(http://www.theplantlist.org/)。利用在线软件phylomatic[31](http://phylodiversity.net/phylomatic/)构建谱系树,将样地调查所得物种名录及其科属信息输入在线软件phylomatic中,自动生成基于被子植物分类系统APGIII且具有进化枝长度的谱系树结构(phylomatic tree, version: Zanne 2014)[32-33],Zanne 2014数据库是基于来自GenBank的7个DNA片段(18S rDNA, 26S rDNA, ITS, matK, rbcL, atpB, trnL-F)使用最大似然法构建的宏谱系树,能提供更加精确可靠的物种之间的进化信息[33]。本研究中使用的谱系树未包括裸子植物。
1.4 谱系指数的选择
Faith[26]于 1992年将群落中物种在谱系树上的进化枝长度总和定义为群落谱系多样性指数(phylogenetic diversity,PD),并得到广泛应用。Webb等[14]提出使用净谱系亲缘关系指数(net relatedness index,NRI)和净最近种间亲缘关系指数(net nearest taxa index,NTI)来表示群落谱系结构,二者分别表示经物种丰富度校正过的平均谱系距离(mean phylogenetic distance,MPD)和平均最近种间谱系距离(mean nearest phylogenetic taxa distance,MNTD)的大小。本文选择近年来广泛使用的NRI指数来表示群落谱系结构,其计算结果与NTI基本一致[12],但NRI指数更加侧重于从整体上而非分支末端结构(NTI)来衡量群落谱系结构[15]。计算NRI指数前,首先假设由群落调查所得87种被子植物组成一个局域物种库[34],然后计算每个样地全部物种的MPD,保持每个样地的物种总数不变,从局域物种库中随机抽取等量物种,共重复999次,进而获得该样地在随机零模型下物种MPDnull分布[35]。最后利用随机分布的结果将MPDobs观察值标准化。NRI计算公式为:
式中,MPDobs表示每个样地MPD的实际观测值,mean(MPDnull)表示每个样地999个随机分布MPD的平均值,sd(MPDnull)表示每个样地999个随机分布MPD值的标准偏差。若NRI>0,说明该样地物种在谱系结构上聚集;若NRI<0,说明该样地物种在谱系结构上发散;若NRI=0,说明该样地物种在谱系结构上随机。NRI指数是基于样地中物种存在与否计算的,并未对其进行多度加权。使用R 3.1.3(R Development Core Team 2013)[36]中picante[37]软件包计算PD和NRI指数。
1.5 环境因子
选择年均温度、年均降水量和土壤含水量作为环境变量来检验群落谱系结构与环境因子之间的关系。由于柴达木盆地地区气象站点较少,样地附近实际气象资料缺乏,因此本研究中使用的年均温度、年均降水量数据均来自世界气候数据网站(http://www.worldclim.org/),该数据库基于1950~2000年世界范围内的气候站点数据,采用薄板平滑样条算法(the thin-plate smoothing spline algorithm)产生[38]。利用每个样点的经纬度数据通过ArcGIS软件提取每个样地的月均温度和月均降雨量信息,进而计算出年均温度和年均降水量[39]。土壤水分由土壤水分仪(TDR)测定。
1.6 数据分析
本研究主要进行了3种分析:(1)使用一般线性模型分析群落谱系结构与环境变量之间的关系。(2)根据灌丛生境差异,将这20个样地的灌丛分为典型荒漠生境灌丛和河谷(河漫滩)生境灌丛,在95%的置信区间内,使用双样本t检验分析河谷(河漫滩)生境灌丛与典型荒漠生境灌丛群落谱系结构及谱系多样性的差异。(3)随后,利用方差分解的方法来检验年均温度、年均降水量及土壤含水量对群落谱系结构独立及交互贡献率[40]。使R 3.1.3(R Development Core Team 2013)[36]进行数据分析,方差分解图使用R进行绘制,其余图均使用Excel进行绘制。
2 结果与分析
2.1 群落谱系结构与年均温度、降水及土壤含水量之间的关系
由表1可以看出,本研究调查的柴达木盆地20个灌丛样地中,有14个样地(70%)的灌丛群落净谱系亲缘关系指数NRI大于0,表明这些群落是由亲缘关系紧密的物种构成;另外有6个样地(30%)的灌丛群落NRI指数小于0,表明构成这些群落的物种亲缘关系较疏远。由此可见,相对于中性理论,基于生态位理论的生境过滤和种间竞争在群落构建过程中可能发挥着更为重要的作用。
通过群落谱系结构与年均降水量和年均温度的回归分析发现,群落谱系结构与年均降水量之间无明显相关关系(NRI:R2=0.05,P=0.77)(图1,A),同样随着年均温度的增加群落谱系结构也没有表现出显著的变化趋势(NRI:R2=0.01,P=0.40)(图1,B)。群落谱系结构与土壤含水量之间存在极显著相关性(NRI:R2=0.69,P=2.77×10-6),随着土壤含水量的增加群落谱系结构发散程度逐渐增大(图1,C)。根据灌丛生境差异,将这20个样地分为河谷(河漫滩)生境和典型荒漠生境,2种生境土壤含水量相差很大。经分析发现,生长在河谷(河漫滩)生境灌丛群落的谱系结构和生长在典型荒漠生境的灌丛群落谱系结构之间存在显著差异(P<0.05)(图2,A),整体上河谷(河漫滩)生境灌丛的谱系结构聚集程度显著低于典型荒漠生境灌丛,说明构成河谷(河漫滩)生境灌丛的物种之间亲缘关系较构成典型荒漠生境灌丛物种的疏远;不同生境灌丛群落的谱系多样性指数(PD)也存在显著差异(P<0.05)(图2,B)。群落谱系多样性指数(PD)与土壤含水量之间也存在极显著相关关系(PD:R2=0.27,P=0.01)(图3)。通过对柴达木盆地年均降水和年均温度做相关性分析,发现年均降水量和年均温度之间存在极显著的负相关关系(R2=0.52,P=1.96×10-4)(图1,D),说明柴达木地区暖干化程度十分明显。
2.2 年均温度、年均降水量及土壤含水量对群落净谱系亲缘关系指数的贡献率
不同生境条件下灌丛群落谱系结构表现出显著性差异。通过方差分解(图4)发现,土壤含水量对群落谱系结构指数NRI的贡献率最大,高达72.47%,土壤含水量和年均温度之间的交互影响对群落谱系结构指数NRI的贡献率为1.01%,年均降水量和年均温度的交互对群落谱系结构指数NRI的贡献率为0.39%。
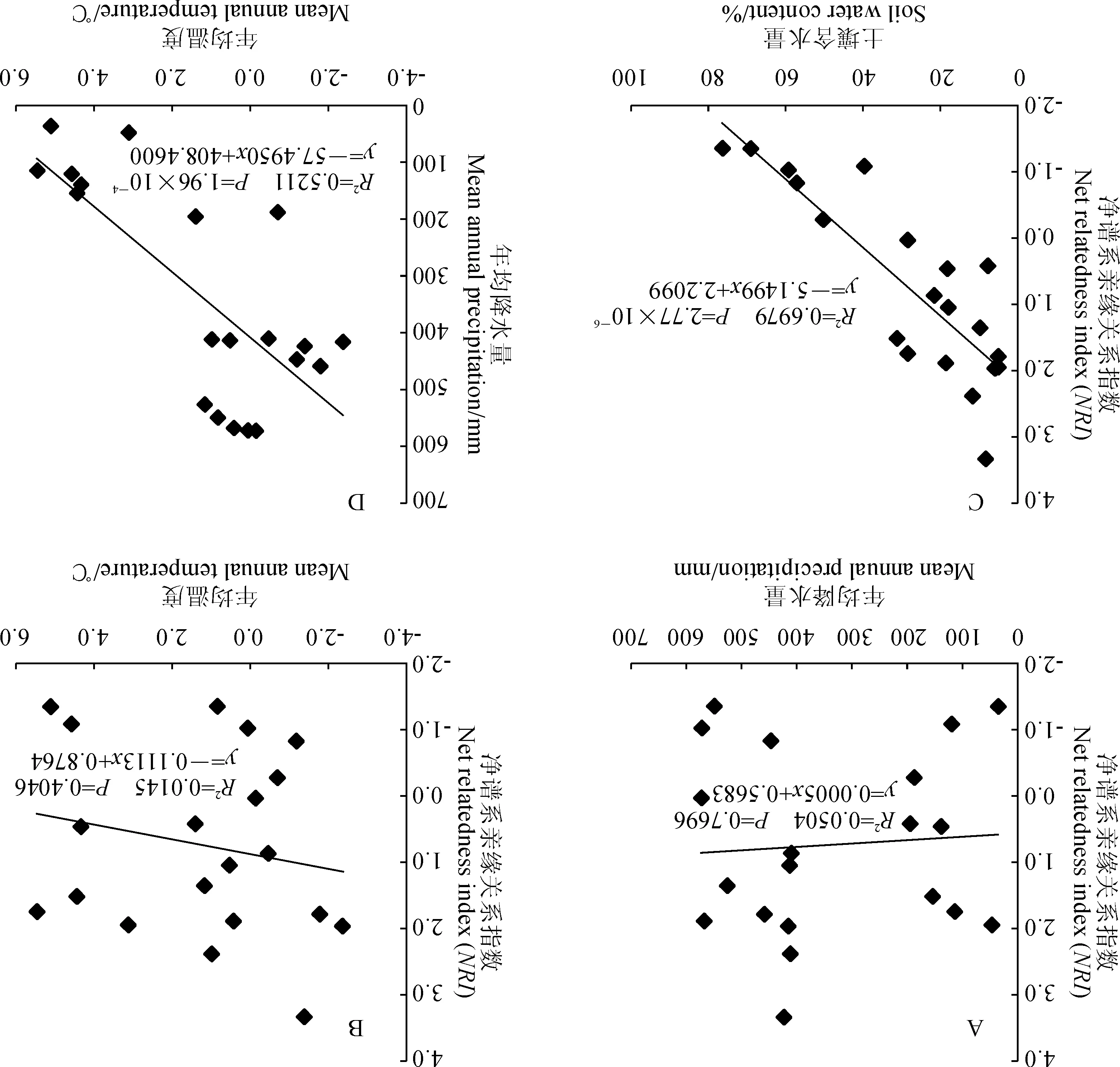
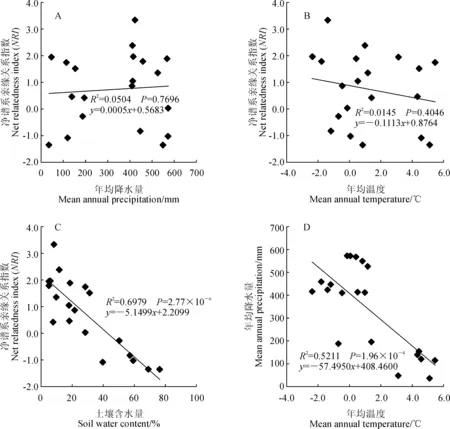
图1 净谱系亲缘关系指数(NRI)和年均降雨量(MAP)、年均温度(MAT)、土壤含水量(SWC),以及年均降雨量(MAP)和年均温度(MAT)散点图Fig.1 Plots of NRI and mean annual precipitation (MAP), mean annual temperature (MAT), soil water content (SWC), and plots of MAP and MAT
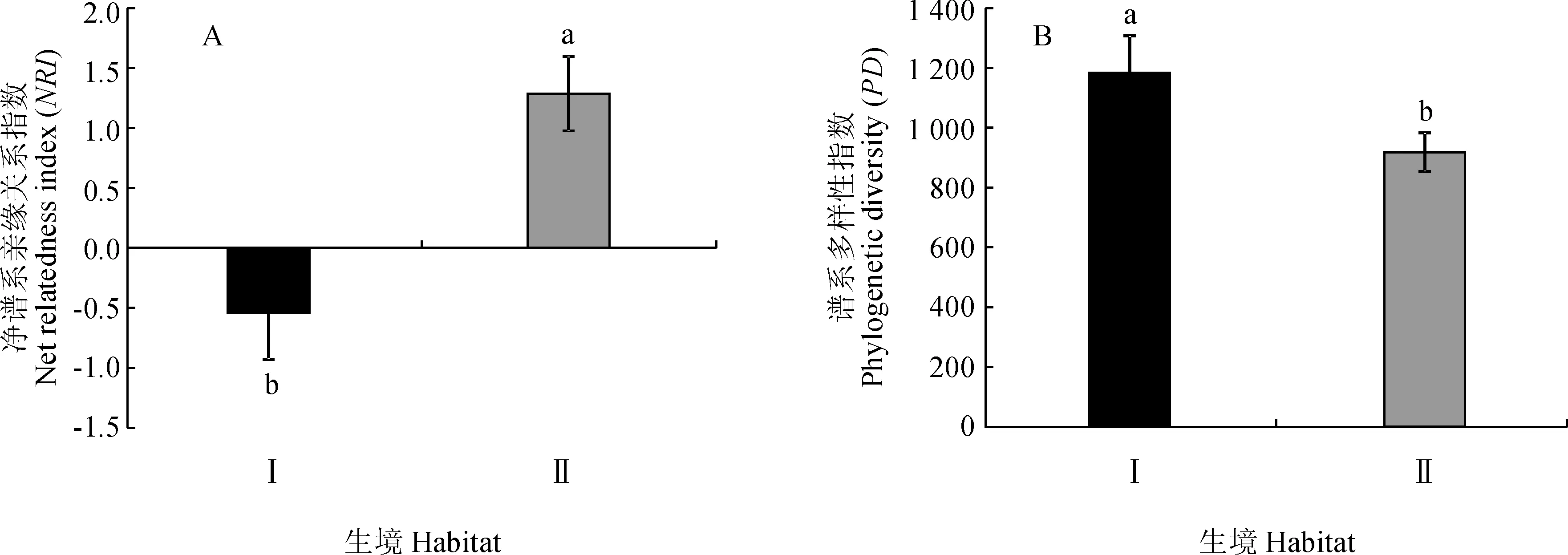
Ⅰ.河谷(河漫滩)生境灌丛;Ⅱ.典型荒漠生境灌丛;不同小写字母表示0.05水平差异显著图2 不同生境灌丛群落系统发育多样性指数(PD)和净谱系亲缘关系指数(NRI)Ⅰ. Shrubs in river valley (river floodplain) habitat;Ⅱ. Shrubs in typical desert habitat; Different normal letters mean significant difference at 0.05 levelFig.2 PD and NRI of shrub communities in different habitats
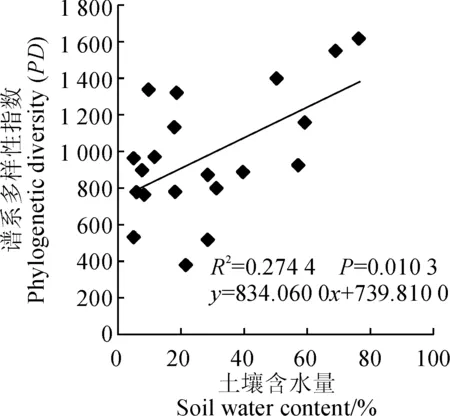
图3 群落谱系多样性指数(PD)与土壤含水量(SWC)散点图Fig.3 Plot of Phylogenetic diversity (PD) and Soil water content (SWC)
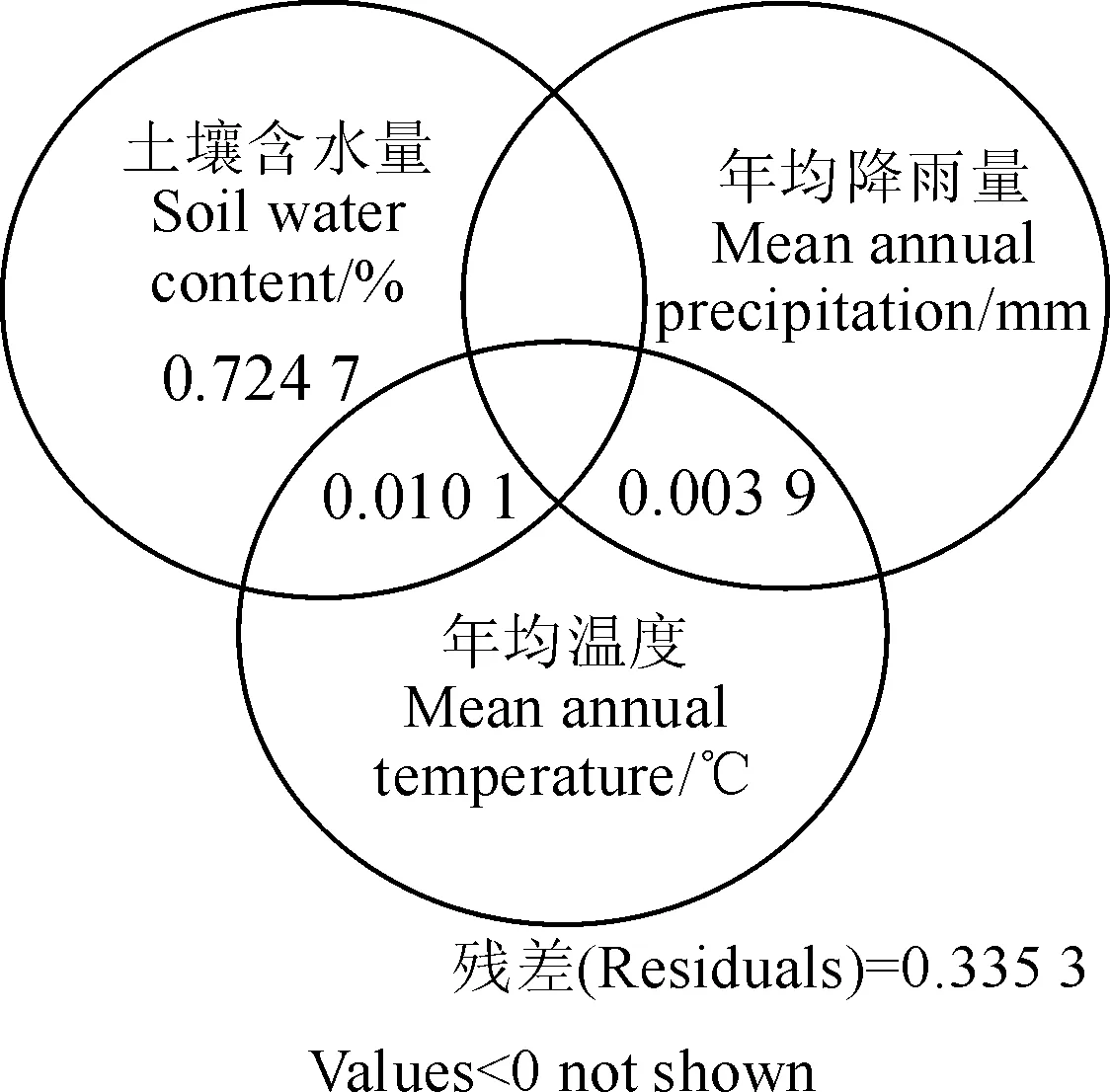
图4 净谱系亲缘关系指数在土壤含水量(SWC)、年均降雨量(MAP)和年均温度(MAT)间的方差分解Fig.4 Variation partitioning of NRI into the independent effects of soil water content (SWC), mean annual precipitation (MAP) and mean annual temperature (MAT), and their overlaps
3 讨 论
群落物种组成是进化历史和生态过程共同作用的结果,研究物种间谱系关系可以使我们在物种进化的角度上对群落构建机制及其影响因素获得更为深刻的理解[41]。Webb等[11]于2000年首次将群落谱系的概念运用在群落生态学研究中并于2002年[14]系统地阐述了通过比较群落内物种之间的谱系距离与随机零模型下谱系距离是否有显著差异来量化群落中物种多样性维持机制即群落构建机制,提出了3种谱系结构(聚集谱系结构、发散谱系结构、随机谱系结构)[14]。在群落系统发育保守性(物种亲缘关系越近,生态特征越相似)的前提下[42],如果生境过滤主导群落构建,环境压力将选择具有相似适应特征、亲缘关系相近的物种共存,群落谱系结构表现为聚集状态;如果亲缘关系较近且具有相似资源利用方式的物种相互竞争或者亲缘关系较疏远物种之间的相互促进作用主导群落构建,则群落中物种之间的亲缘关系疏远,谱系结构表现为发散状态;如果中性过程在群落构建过程中发挥主导作用,则群落呈现出随机的谱系结构[14]。还有研究发现,随机的谱系结构也可能是生境过滤和种间竞争共同作用的结果[43]。此后,越来越多的学者将群落谱系结构的观点应用在群落构建机制的探索以及生物多样性保护中[14,16-17,20,25,31]。
3.1 柴达木盆地灌丛群落构建机制
通过结果分析可知,柴达木盆地灌丛70%的群落净谱系亲缘关系指数NRI>0,30%的群落净谱系亲缘关系指数NRI<0,即大多数群落表现出聚集的谱系结构,这与中性理论所预测的物种分布趋向于随机,呈现随机谱系结构的结论是不相符的[14]。结果表明在柴达木盆地多数灌丛群落由亲缘关系紧密的物种组成,结合柴达木盆地严苛的气候特点,推测生境过滤作用在大多数群落构建中可能发挥主导作用,说明相对于中性理论来说生态位理论在柴达木盆地灌丛群落构建过程中可能具有更加重要的作用。
研究群落谱系结构与环境因子的关系有助于我们对群落构建机制的理解。水分条件是半干旱、干旱生态系统最主要的限制因子[44],柴达木盆地作为极端干旱的生态系统[28],物种面临干旱的环境压力可能会迫使亲缘关系相近、具有相似抗旱能力的物种共存,干旱的环境压力可能会随着降雨量的增加而减弱。Li 等[45]在横断山区研究发现,随着年均降雨量的增加,NRI指数聚集程度整体上呈现出减小的趋势,表明年均降雨量的增加使环境条件更加优越,群落中亲缘关系较远的物种得以共存。但是在本研究中净谱系亲缘关系指数NRI随着年均降雨量的增加并没有表现出显著的变化趋势。造成这种结果的原因可能是柴达木盆地年均蒸发量远远大于降雨量[28],从而降低甚至完全抵消降雨对灌丛群落生长和局域环境条件优化的促进效应,强烈的环境过滤作用会筛选出具有相似抗旱能力的物种共存。Qin[46]等在黄土高原东部油松(Pinustabuliformis)林中发现无论是在吕梁山还是太行山植物群落净谱系亲缘关系指数NRI均与降雨不存在相关性,这一结果在一定程度上支持了我们的结果和推断。群落净谱系亲缘关系指数NRI与年均温度之间也不存在显著的相关性。这可能是因为水分是柴达木盆地最主要的限制因子[44],根据限制性因子定律,在水分限制没有得到明显缓解之前,灌丛群落的物种组成只与有效的水分含量有关。柴达木盆地暖干化趋势很明显,也就是说随着温度的升高进一步加剧了降雨因子的限制性作用。
柴达木盆地灌丛群落净谱系亲缘关系指数NRI与土壤含水量之间存在极显著相关关系(R2=0.69,P=2.77×10-6),随着土壤含水量的增加群落谱系结构表现出由聚集向发散过渡的趋势,构成群落的物种之间亲缘关系紧密程度逐渐减小。灌丛群落所处的具体生境可能是导致土壤含水量差异的主要原因,结合具体生境的土壤类型—土壤含水量信息我们发现,从风沙土、灰棕漠土、棕钙土、沼泽土到草甸土,土壤含水量总体上由11.70%、12.65%、31.48%、52.08%上升到72.74%,群落净谱系亲缘关系指数NRI也由1.68、1.43、0.98、-0.98过渡到-1.35,即随着生境含水量的增加,群落谱系结构发散程度逐渐增大。土壤水分是荒漠植被水分的主要来源[47],在土壤含水量较低的生境中,环境条件严峻,干旱胁迫较大[48],强烈的生境压力驱使具有相似生理适应特性的物种即亲缘关系较近的物种共存,群落表现出聚集的谱系结构。在土壤含水量较高的生境中,环境和资源条件优越,生态胁迫不明显[49],群落中物种之间的竞争作用可能起主导作用,使亲缘关系较远的物种共存而表现出发散的谱系结构。因此,在高土壤含水量和低土壤含水量生境的群落构建过程中生态位理论可能发挥主导作用。在土壤含水量适中的生境中,部分群落的净谱系亲缘关系指数NRI尽管不等于0,但无显著统计学差异,这可能是种间竞争和严苛的环境压力相互作用的结果[43]。有研究表明根系吸水位层相似的植物之间存在竞争[50],不同物种根深及吸水深度存在差异[51]。在柴达木盆地土壤水分适中的环境中,亲缘关系紧密的物种可能具有相近的地下生态位,亲缘关系较远的物种可能因土壤水分利用生态位具有较大差异而得以共存。严苛的环境压力(如冬季低温)可能会抵消种间竞争的效应进而形成随机的谱系结构[43],是否中性理论在群落构建过程中也发挥重要作用则需要结合植物功能性状在未来作进一步研究。方差分解的结果显示,土壤含水量对净谱系亲缘关系指数NRI有最高的贡献率,从另一个侧面再次说明了土壤含水量与NRI之间的紧密联系。从低土壤含水量生境中的谱系结构聚集到高土壤含水量生境中的谱系结构发散表现出生境过滤作用逐步减弱,生物间相互作用逐渐加强的变化趋势。
3.2 柴达木盆地灌丛群落谱系结构在生物多样性保护中的应用
在全球变化的背景下,生物多样性正面临前所未有的风险与挑战,生物多样性丧失的情形日益严重[1,2]。尤其是在干旱生态系统,现存生物多样性消失后再恢复几乎是不可能的。因此科学有效的生物多样性保护策略显得尤为重要。以往的生物多样性保护观点认为所有物种具有相同的保护价值[52],然而,有研究证明不同的物种因进化历史的差异而具有不同的保护价值[20],以往仅基于物种多样性水平制定的保护策略是不全面的[53],有效的保护策略应充分考虑物种进化水平的谱系多样性[4,20,23-24]。Cadotte等[54]研究发现谱系多样性高的群落具有更高的稳定性,Pu等[22]发现较高的谱系多样性促进群落初级生产的稳定性,更重要的是,谱系多样性较高的群落在未来面对全球气候变化时拥有更强的进化潜力[23]。据此,在采取保护行动时应该优先保护谱系多样性高的群落。在本研究中发现随着土壤含水量的增加,柴达木盆地灌丛群落谱系多样性指数(PD)表现出了显著升高的趋势,较典型荒漠生境,河谷(河漫滩)生境中的灌丛群落拥有更高的谱系多样性,基于先前的研究结果,我们认为在柴达木盆地生物多样性保护实际行动中应优先保护河谷(河漫滩)生境的灌丛群落。Sechrest等[55]研究发现谱系多样性丧失速率远远快于物种多样性,所以优先保护河谷(河漫滩)生境谱系多样性高的灌丛群落就显得更加迫切重要。Forest等[56]在研究植物物种多样性和谱系多样性分布格局时,发现谱系结构发散的群落,其谱系多样性要高于估计值;而谱系结构聚集的群落,其谱系多样性可能要低于估计值。本研究发现河谷(河漫滩)生境灌丛群落谱系结构的聚集程度要远远低于典型荒漠生境灌丛群落,因此河谷(河漫滩)生境的灌丛群落则可能拥有比估计值更高的谱系多样性,优先保护这些群落可能为将来物种演化保留更大的潜力。除此之外,河谷(河漫滩)生境灌丛在柴达木盆地水源涵养,局域小气候改善方面发挥着更为直接的作用[57],对其进行优先保护对于柴达木地区未来的可持续发展具有更为重要的现实意义。
本研究是群落谱系的研究方法在青藏高原干旱荒漠生态系统的首次应用,初步回答了荒漠生态系统不同生境下灌丛群落的谱系结构、主导群落构建的主要生态过程及与部分环境因子的关系,并在群落谱系的角度为柴达木盆地保育生物学研究提供了理论基础。在未来的研究中需要进一步将植物功能性状、更加全面的环境因子与高分辨率的谱系树结合起来对柴达木盆地等干旱荒漠生态系统及其他生态系统进行群落谱系研究,以期获得更科学、深入的群落构建机制,为生物多样性保护行动提供更加可靠的理论基础。
参考文献:
[1] THOMAS J A, TELFER M G, ROY D B,etal. Comparative losses of british butterflies, birds, and plants and the global extinction crisis[J].Science, 2004, 303: 1 879-1 881.
[2] THUILLER W, LAVERGNE S, ROQUET C,etal. Consequences of climate change on the tree of life in Europe[J].Nature, 2011,470(7 335): 531-534.
[3] CARDINALE B J, DUFFY J E, GONZALEZ A,etal. Biodiversity loss and its impact on humanity[J].Nature, 2012,486(7 401):59-67.
[4] 马克平. 生物多样性科学的热点问题[J]. 生物多样性, 2016,24(1): 1-2.
MA K P. Hot topics for Biodiversity Science[J].BiodiversityScience, 2016,24(1): 1-2.
[5] 马克平. 第19届国际植物学大会盛况空前,生物多样性科学受到特别关注[J]. 生物多样性, 2017,25(8): 797-798.
MA K P. The best case in the history of International Botanical Congress and biodiversity science receiving special attention in the 19th IBC[J].BiodiversityScience, 2017,25(8): 797-798.
[6] 于伯华, 吕昌河. 青藏高原高寒区生态脆弱性评价[J]. 地理研究, 2011,30(12): 2 289-2 295.
YU B H, LÜ C H. Assessment of ecological vulnerability on the Tibetan Plateau[J].GeographicalResearch, 2011,30(12): 2 289-2 295.
[7] 杨荣金, 舒俭民, 李秀红,等. 柴达木盆地生态环境保护战略与对策[J]. 科技导报, 2017,35(6): 115-119.
YANG R J, SHU J M, LI X H,etal. Strategy and countermeasures of ecological environment protection in Qaidam Basin[J].ScienceandTechnologyReview, 2017,35(6): 115-119.
[8] 柴永福, 岳 明. 植物群落构建机制研究进展[J]. 生态学报, 2016,36(15): 4 557-4 572.
CHAI Y F,YUE M.Research advances in plant community assembly mechanisms[J].ActaEcologicaSinica,2016,36(15): 4 557-4 572.
[9] GÖTZENBERGER L, BELLO F D, BRÅTHEN K A,etal. Ecological assembly rules in plant communities—approaches, patterns and prospects[J].BiologicalReviewsoftheCambridgePhilosophicalSociety, 2012,87(1): 111-127.
[10] VELLEND M. Conceptual synthesis in community ecology[J].QuarterlyReviewofBiology, 2010,85(2): 183-206.
[11] WEBB C O. Exploring the phylogenetic structure of ecological communities: An example for rain forest trees[J].AmericanNaturalist, 2000,156(2): 145-155.
[12] SWENSON N G, ENQUIST B J, THOMPSON J,etal. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities[J].Ecology, 2007,88(7): 1 770-1 780.
[13] SWENSON N G, STEGEN J C, DAVIES S J,etal. Temporal turnover in the composition of tropical tree communities: Functional determinism and phylogenetic stochasticity[J].Ecology, 2012, 93: 490-499.
[14] WEBB C O,ACKERLY D D,MCPEEK M A,etal. Phylogenies and community ecology[J].AnnualReviewofEcologyandSystematics, 2002.33: 475-505.
[15] LETCHER S G. Phylogenetic structure of angiosperm communities during tropical forest succession[J].ProceedingsBiologicalSciences, 2010,277(1 678): 97-104.
[16] 姜晓燕, 梁林峰, 毕润成,等. 山西霍山植物群落谱系结构的空间格局[J]. 西北植物学报, 2016,36(12): 2 505-2 512.
JIANG X Y, LIANG L F, BI R C,etal. Spatial pattern of phylogenetic structure of plant community in Shanxi Huoshan Mountain[J].ActaBotanicaBoreali-OccidentaliaSinica, 2016,36(12): 2 505-2 512.
[17] HUANG J X, ZHANG J, SHEN Y,etal. Different relationships between temporal phylogenetic turnover and phylogenetic similarity and in two forests were detected by a new null model[J].PlosOne, 2014,9(4): e95703.
[18] 葛学军. DNA条形码在植物系统发育区系学研究中的应用[J]. 生物多样性, 2015,23(3): 295-296.
GE X J. Application of DNA barcoding in phylofloristics study[J].BiodiversityScience, 2015,23(3): 295-296.
[19] WINTER M, DEVICTOR V, SCHWEIGER O. Phylogenetic diversity and nature conservation: where are we?[J].TrendsinEcology&Evolution, 2013,28(4): 199-204.
[20] LAITY T, LAFFAN S W, GONZLEZ-OROZCO C E,etal. Phylodiversity to inform conservation policy: An Australian example[J].ScienceoftheTotalEnvironment, 2015, 534: 131-143.
[21] FLYNN D F, MIROTCHNICK N, JAIN M,etal. Functional and phylogenetic diversity as predictors of biodiversity-ecosystem-function relationships[J].Ecology, 2011,92(8): 1 573-1 581.
[22] PU Z, DAYA P, TAN J,etal. Phylogenetic diversity stabilizes community biomass[J].JournalofPlantEcology, 2014,7(2): 176-187.
[23] 慈秀芹, 李 捷. 系统发育多样性在植物区系研究与生物多样性保护中的应用[J]. 生物多样性, 2017,25(2): 175-181.
CI X Q, LI J. Phylogenetic diversity and its application in floristics and biodiversity conservation[J].BiodiversityScience, 2017,25(2): 175-181.
[24] GEO BON. Adequacy of Biodiversity Observation Systems to support the CBD 2020 Targets.AreportpreparedbytheGrouponEarthObservationsBiodiversityObservationNetwork(GEOBON)fortheConventiononBiologicalDiversity.GEOBON,Pretoria2011: 1-106.
[25] DEVICTOR V, MOUILLOT D, MEYNARD C,etal. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: the need for integrative conservation strategies in a changing world[J].EcologyLetters, 2010,13: 1 030-1 040.
[26] FAITH D P. Conservation evaluation and phylogenetic diversity[J].BiologicalConservation, 1992,61(1): 1-10.
[27] 朱文彬, 吕爱锋, 贾绍凤. 基于NDVI的柴达木盆地植被空间分异规律及影响因素[J]. 干旱区研究, 2010,27(5): 691-698.
ZHU W B, LÜ A F, JIA S F. Study on spatial distribution of vegetation coverage and its affecting factors in the Qaidam Basin based on NDVI[J].AridZoneResearch, 2010,27(5): 691-698.
[28] 刘燕华. 柴达木盆地水资源合理利用与生态环境保护[M]. 北京:科学出版社, 2000.
[29] 于海洋, 张振德, 张佩民. 青海土地荒漠化评价及动态监测[J]. 干旱区研究, 2007,24(2): 153-158.
YU H Y, ZHANG Z D, ZHANG P M. RS- and GIS-based evaluation and dynamic monitoring of land desertification in Qinghai Province[J].AridZoneResearch, 2007,24(2): 153-158.
[30] 生态系统固碳项目技术规范编写组. 生态系统固碳观测与调查技术规范[M]. 北京:科学出版社, 2015.
[31] WEBB C O, DONOGHUE M J. Phylomatic: tree assembly for applied phylogenetics[J].MolecularEcologyNotes, 2005, 5: 181-183.
[32] ZANNE A E, AL E. Three keys to the radiation of angiosperms into freezing environments.[J].Nature, 2014,506(7 486): 89-92.
[33] BARAK R S, WILLIAMS E W, HIPP A L,etal. Restored tallgrass prairies have reduced phylogenetic diversity compared with remnants[J].JournalofAppliedEcology, 2017, (54): 1 080-1 090.
[34] 方精云, 王襄平, 唐志尧. 局域和区域过程共同控制着群落的物种多样性:种库假说[J]. 生物多样性, 2009,17(6): 605-612.
FANG J Y, WANG X P, TANG Z Y. Local and regional processes control species richness of plant communities: the species pool hypothesis[J].BiodiversityScience, 2009,17(6): 605-612.
[35] SWENSON N G, ANGLADA-CORDERO P, BARONE J A. Deterministic tropical tree community turnover: evidence from patterns of functional beta diversity along an elevational gradient[J].ProceedingsBiologicalSciences, 2011,278(1 707): 877-884.
[36] R DEVELOPMENT CORE TEAMR:ALanguageandEnvironmentforStatisticalComputing. R Foundation for Statistical Computing[M],Vienna, Austria, 2013.
[37] KEMBEL S W, COWAN P D, HELMUS M R,etal. Picante: R tools for integrating phylogenies and ecology[J].Bioinformatics, 2010,26: 1 463-1 464.
[38] HIJMANS R J, CAMERON S E, PARRA J L,etal. Very high resolution interpolated climate surfaces for global land areas[J].InternationalJournalofClimatology, 2005,25(15): 1 965-1 978.
[39] 田 聪, 穆尼热, 朱忠艳,等. 基于DIVA-GIS快速获取气候信息的方法[J]. 农学学报, 2015,5(5): 109-113.
TIAN C, MU N R, ZHU Z Y,etal. Method for the rapid obtaining of climate data based on DIVA-GIS[J].JournalofAgriculture, 2015,5(5): 109-113.
[40] LI X H, SUN H. Phylogenetic pattern of alpine plants along latitude and longitude in Hengduan Mountains Region[J].PlantDiversity, 2017,39(1): 37-43.
[41] 黄建雄, 郑凤英, 米湘成. 不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响[J]. 植物生态学报, 2010,34(3): 309-315.
HUANG J X, ZHENG F Y, MI X C. Influence of environmental factors on phylogenetic structure at multiple spatial scales in an evergreen broad-leaved forest of China[J].ChineseJournalofPlantEcology, 2010,34(3): 309-315.
[42] BURNS J H, SCHMITT J. More closely related species are more ecologically similar in an experimental test[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 2011,108(13): 5 302-5 307.
[43] HELMUS M R, SAVAGE K, DIEBEL M W,etal. Separating the determinants of phylogenetic community structure[J].EcologyLetters, 2007,10(10): 917-925.
[44] HARPOLE W, POTTS D, SUDING K. Ecosystem responses to water and nitrogen amendment in a California grassland[J].GlobalChangeBiology, 2007,13(11): 2 341-2 348.
[45] XIN-HUI LI, ZHU X X, NIU Y,etal. Phylogenetic clustering and overdispersion for alpine plants along elevational gradient in the Hengduan Mountains Region, southwest China[J].JournalofSystematicsandEvolution, 2014,52(3): 280-288.
[46] QIN H, DONG G, ZHANG Y,etal. Patterns of species and phylogenetic diversity ofPinustabuliformisforests in the eastern Loess Plateau, China[J].ForestEcology&Management, 2017, 394: 42-51.
[47] SNYDER K A, WILLIAMS D G. Water sources used by riparian trees varies among stream types on the San Pedro River, Arizona.[J].Agricultural&ForestMeteorology, 2000,105(1): 227-240.
[48] WILCOX C S, FERGUSON J W, FERNANDEZ G C J,etal. Fine root growth dynamics of four Mojave Desert shrubs as related to soil moisture and microsite[J].JournalofAridEnvironments, 2004, 56: 129-148.
[49] 王家强, 韩 路, 柳维扬, 等.塔里木河中游荒漠绿洲过渡带土壤水分与植被空间格局变化关系研究[J].西北林学院学报,2018,33(1): 1-10.
WANG J Q, HAN L, LIU W Y,etal. Variable relationship of soil moisture and spatial pattern along desert oasis transition zone in the Tarim River middle reaches basin[J].JournalofNorthwestForestryUniversity, 2018,33(1): 1-10.
[50] 曾 巧, 马剑英. 黑河流域不同生境植物水分来源及环境指示意义[J]. 冰川冻土, 2013,35(1): 148-155.
ZENG Q, MA J Y. Plant water sources of different habitats and its environmental indication in Heihe River basin[J].JournalofGlaciologyandGeocryology, 2013,35(1): 148-155.
[51] 徐贵青, 李 彦. 共生条件下三种荒漠灌木的根系分布特征及其对降水的响应[J]. 生态学报, 2009,29(1): 130-137.
XU G Q, LI Y. Roots distribution of three desert shrubs and their response to precipitation under co-occurring conditions[J].ActaEcologicaSinica, 2009,29(1): 130-137.
[52] POSADAS P, ESQUIVEL D R M, CRISCI J V. Using phylogenetic diversity measures to set priorities in conservation: an example from southern South America[J].ConservationBiology, 2001,15(5): 1 325-1 334.
[53] LIN W, YOUNG S S, WEI W,etal. Conservation priorities of forest ecosystems with evaluations of connectivity and future threats: Implications in the Eastern Himalaya of China[J].BiologicalConservation, 2016,195(2 016): 128-135.
[54] CADOTTE M W, DINNAGE R, TILMAN D. Phylogenetic diversity promotes ecosystem stability[J].Ecology, 2012,93(8): S223-S233.
[55] SECHREST W, BROOKS T M, FONSECA G A B D,etal. Hotspots and the conservation of evolutionary history[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 2002,99(4): 2 067-2 071.
[56] FOREST F, GRENYER R, ROUGET M,etal. Preserving the evolutionary potential of floras in biodiversity hotspots[J].Nature, 2007,445(7 129): 757-60.
[57] 赵 钧. 柴达木盆地林业生态建设探讨[J]. 青海农林科技, 2007, (4): 58-60.
ZHAO J. Approach on construction forestry ecology in Chaidamu Basin[J].ScienceandTechnologyofQinghaiAgricultureandForestry, 2007, (4): 58-60.
猜你喜欢
作物学报(2022年2期)2022-11-06 12:11:02
水土保持学报(2022年5期)2022-10-10 14:20:06
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01 11:26:42
中国现代中药(2021年9期)2021-11-16 07:43:32
矿产勘查(2020年6期)2020-12-25 02:41:08
沉积与特提斯地质(2019年4期)2019-07-19 06:04:20
草业科学(2019年5期)2019-06-04 09:36:24
天然产物研究与开发(2018年10期)2018-11-06 07:43:46
广东农业科学(2017年5期)2017-08-29 10:37:26
天津师范大学学报(自然科学版)(2016年4期)2016-12-14 06:04:26
