
干旱胁迫对小紫珠光合生理的影响
2018-06-12 06:55杨建伟孙桂芳刘嘉翔史宝胜
西北植物学报 2018年4期
杨建伟,赵 丹,孙桂芳,刘嘉翔,安 彤,史宝胜
(河北农业大学 园林与旅游学院,河北保定 071000)
中国是一个水资源相对贫乏的国家,人均水资源占有量为世界平均水平的1/4[1-2]。中国的华北、西北、内蒙古和青藏高原绝大部分地区属于干旱、半干旱地区[3],而且干旱的趋势还在不断的加剧。干旱对植物的正常生长产生严重影响,成为限制植物生存的重要环境因素[4]。光合作用是植物生长发育和生物量积累的基础,干旱胁迫会影响植物对光能的消耗,导致大量的光能过剩,损伤植物的光合器官。柴胜丰等[5]对干旱胁迫下金花茶光合生理进行了研究,结果表明随着干旱胁迫的加剧植物的光合作用逐渐下降,在严重干旱胁迫下引发了光合机构的损伤和光合电子的传递。郭有燕等[6]的研究发现,干旱条件下黑果枸杞的初始荧光(Fo)和非光化学淬灭系数qN逐渐增加,而其最大荧光(Fm)、PSⅡ最大光化学效率(Fv/Fm)和光化学淬灭系数(qP)均逐渐降低。干旱胁迫对植物的光合作用产生严重的影响,因此研究不同水分条件对植物光合生理的影响能够深入了解干旱对植物伤害的内在机制。
小紫珠(Callicarpadichotoma)属马鞭草科紫珠属,夏季枝叶繁茂,入秋紫果累累,经冬不落,极具观赏价值[7]。目前对于小紫珠的研究大多集中在栽培技术、引种繁殖方面[8],而关于干旱胁迫对小紫珠光合生理的影响尚未见报道。本研究采用人工控制水分条件,探讨不同水分条件对小紫珠光合参数的影响,旨在为小紫珠的园林栽培管理提供理论依据。
1 材料和方法
1.1 试验区概况
试验在河北省保定市(38°10′~40°00′ N,113°40′~116°20′ E)河北农业大学标本园进行,该地海拔18 m,年平均气温12.7 ℃,年均降水量575.4 mm,无霜期200 d。四季分明,属暖温带亚湿润气候区。
1.2 试验设计
试验材料为2年生的小紫珠(Callicarpadichotoma),盆栽直径30 cm,高28 cm。盆土为苗圃地园土、砂和营养土自然条件下风干后按照2∶2∶1均匀混合装盆,每盆盆土重13 kg。
试验在自制人工防雨棚中进行,依据罗桑卓玛等[9]研究共设置4个水分处理,分别为田间最大持水量(45.34%)的95%(对照,CK)、75%(轻度干旱,LD)、55%(中度干旱,MD)和35%(重度干旱,SD)。每处理3次重复,每重复4盆。田间持水量用环刀法测定。2017年4月下旬移栽花盆后进行正常水分管理,6月中旬进行控水处理,试验期间每天18:00称重,补充散失水分,控水处理1个月后进行各项指标的测定。
1.3 指标测定
1.3.1光合日变化的测定采用CIRAS-3便携式光合测定系统测定植物叶片的光合作用相关指标。测定时选取叶位及长势基本一致且无病虫害的叶片,每处理测定9片叶片。参照王建华等[10]方法,选择晴朗少云的天气,从6:00至18:00每隔2 h测定1次。测定的内容主要有叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)和水分利用效率(WUE)。
1.3.2光响应曲线的测定采用CIRAS-3便携式光合测定系统测定光响应曲线。参照胡义等[11]方法,选择晴朗少云的天气,在8: 00~10: 00进行。利用CO2钢瓶将参比室的CO2浓度稳定在380 μmol·mol-1,设定温度为25 ℃。LED光源设定2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、150、100、80、60、40、20和0 μmol·m-2·s-1等梯度,测定不同光照有效辐射下的净光合速率(Pn),3次重复。参照叶子飘[12]的方法计算光补偿点(LCP)、光饱和点(LSP)、暗呼吸速率(Rd)、最大净光合速率(Pnmax)和表观量子效率(AQY)。
1.3.3叶绿素荧光参数的测定采用FP100便携式荧光仪,参照魏晓东等[13]方法,选择晴朗少云的天气于上午8:00~10:00,将叶片用叶片夹进行暗适应30 min后,测定植物叶片的叶绿素荧光参数,包括初始荧光(Fo)、最大荧光(Fm)、PSⅡ最大光化学效率(Fv/Fm)、PSⅡ的量子效率(Fv/Fo);能量分配比率参数:φPo、Ψo、φEo、φDo;PSⅡ比活性参数ABS/RC、TRo/RC、ETo/RC、DIo/RC。每处理测定9片叶片。
1.3.4光合色素含量的测定用80%丙酮提取,按Lichtenthaler[14]方法测定提取液在波长663和646 nm下吸光值。计算公式如下:
叶绿素a含量=(12.21×D663-2.81×D646) ×V/(1 000×W)
叶绿素b含量=(20.13×D646-5.03×D663) ×V/(1 000×W)
叶绿素总含量=D652×V/(34.5×W)
其中,V为浸提液最终体积(mL),W为叶鲜重(g)
1.4 数据处理
采用Excel 2017进行数据处理,通过SPSS 22统计软件进行多重比较。
2 结果与分析
2.1 干旱胁迫对小紫珠光合日进程的影响
如图1所示,干旱胁迫下小紫珠Pn、Gs和Tr总体呈下降的趋势,且变化趋势基本一致。小紫珠Pn、Gs和Tr在CK和LS处理下为“单峰型”,分别在12:00、10:00和12:00达到峰值,之后逐渐下降。在MS和SS处理下呈现“双峰型”,与CK相比峰值显著提前2 h,在中午出现“光合午休”现象。小紫珠Ci日变化在CK、LS和MS处理下整体呈先降低后升高的“V字型”,呈现早晚高午间低的趋势,在SS处理下Ci在中午显著升高,在12:00达到最大,与CK相比增加了13.06%。随着干旱胁迫的加剧小紫珠WUE逐渐下降,在CK处理下最高,在SS处理下最低,就日变化而言整体呈现上午高,下午逐渐降低的趋势,在10:00后迅速降低。
2.2 干旱胁迫对小紫珠光响应参数的影响
如图2所示,小紫珠在CK、LS、MS处理下Pn随着PAR升高逐渐增加,当PAR接近饱和光强时,渐趋平缓而后降低,在SS处理下趋势平缓,无明显变化。不同水分条件下小紫珠的Pn对PAR的响应不同,随着PAR的增大小紫珠在CK和LS处理下Pn显著高于MS和SS处理。利用直角双曲线修正模型拟合不同土壤水分条件下小紫珠光响应曲线并计算出相应参数(表1)。与CK相比在LS、MS和SS处理下小紫珠的LSP、Pnmax、Rd和AQY显著下降,分别降低了0.56%、4.38%、69.17%;5.75%、25.18%、97.98%;55.56%、48.54%、93.57%;24.24%、42.42%、51.52%,LCP显著升高,增加了133.11%、619.29%、568.62%。
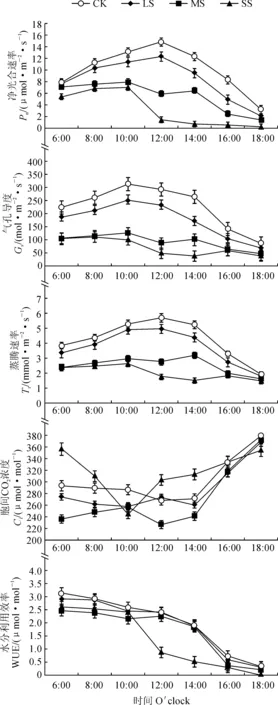
图1 干旱胁迫下小紫珠光合日进程Fig.1 The photosynthetic daily process of the C. dichotoma under drought stress
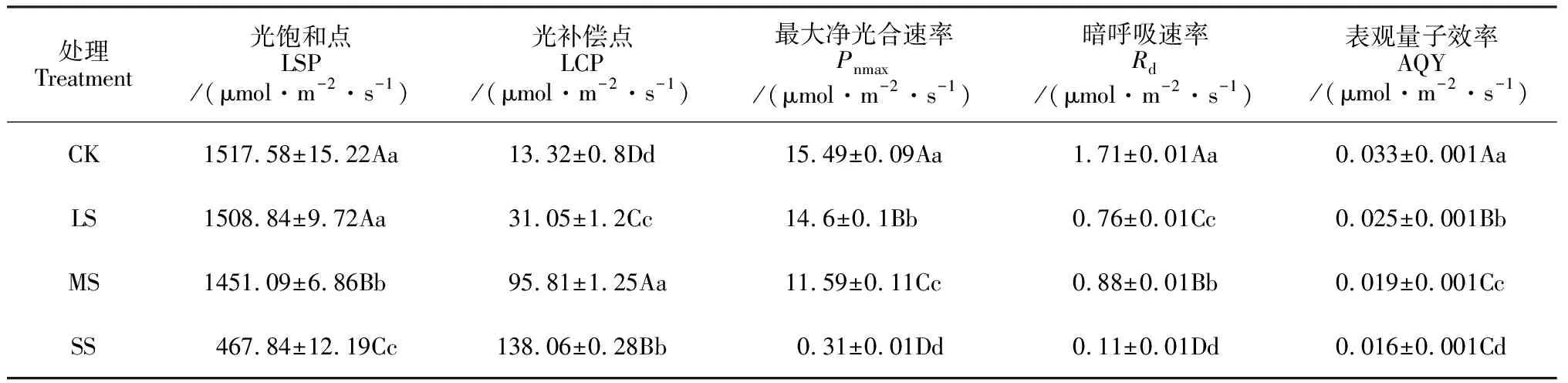
表1 干旱胁迫下小紫珠光响应曲线参数
注:大写字母表示不同处理之间差异达到极显著水平(P<0. 01);小写字母表示不同处理之间差异达到显著水平(P<0.05)。下同
Note:The capital letters indicated the difference between different treatments reached an extremely significant level (P<0.01). The normal letters indicated the difference between different treatments was significant (P<0.05). The same as below
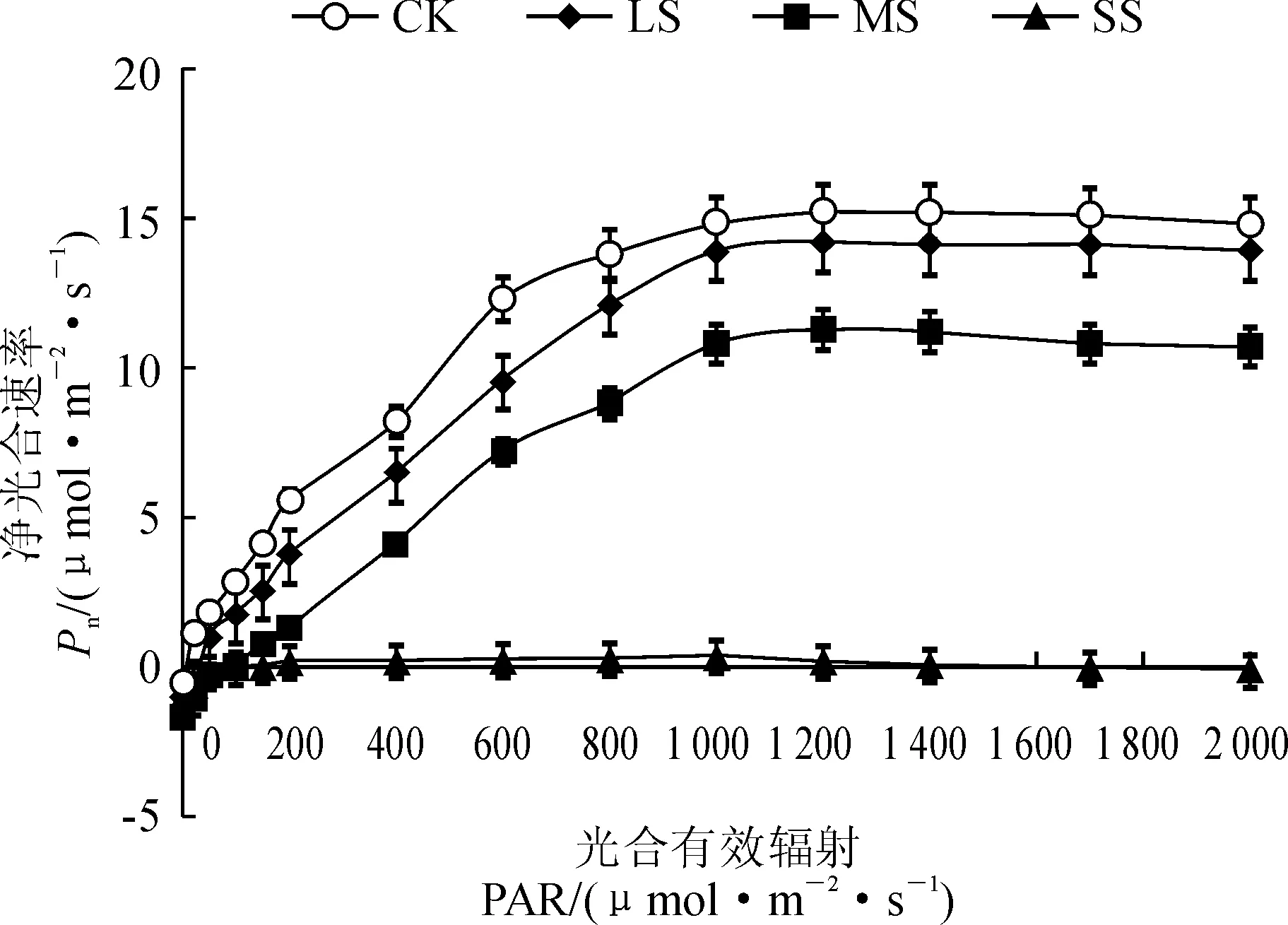
图2 干旱胁迫下小紫珠光响应曲线的变化Fig.2 The light response curve of the C. dichotoma under drought stress
2.3 干旱胁迫对小紫珠叶绿素荧光参数的影响
如图3所示,随着干旱胁迫加剧小紫珠Fo显著上升,在LS、MS、SS处理下与CK相比差异极显著(P<0.01),分别增加了3.66%、11.61%、21.02%。在干旱胁迫下小紫珠Fm逐渐降低,在LS处理下与CK相比差异显著(P<0.05),降低了1.01%,在MS和SS处理下差异极显著(P<0.01),分别降低了7.63%、14.98%。随着胁迫程度增加,小紫珠Fv/Fm逐渐降低,但与CK相比LS和MS处理下并无显著差异(P>0.05),在SS处理下差异极显著(P<0.01),降低了20.99%。随着干旱胁迫加剧,小紫珠Fv/Fo显著降低,在LS、MS、SS处理下与CK相比分别降低了22.09%、25.44%和27.86%。
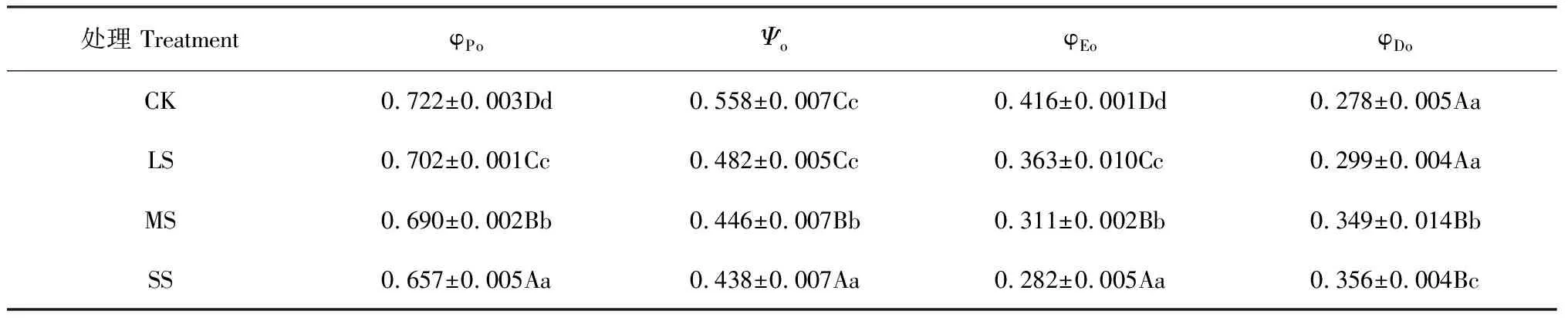
如表2所示,随着干旱胁迫程度的增加,小紫珠φPo显著降低,在LS、MS、SS处理下与CK相比分别减少2.77%、4.43%、9%;与CK相比在LS处理下小紫珠Ψo无显著差异(P>0.05),在MS和SS处理下差异极显著(P<0.01),分别降低了20.07%、21.51%;随着干旱胁迫的增加小紫珠的φEo显著降低,在LS、MS、SS处理下与CK相比分别降低了12.74%、25.24%、32.21%;在LS处理下小紫珠φDo与CK相比无显著差异(P>0.05),在MS和SS处理下差异显著,分别增加了25.54%、28.06%。
如表3所示,随着干旱胁迫程度的增加,小紫珠ABS/RC逐渐上升,与CK相比在MS和SS处理下差异极显著(P<0.01),分别增加了10.99%、33.84%;随着干旱胁迫的加剧小紫珠TRo/RC逐渐升高,各干旱处理与CK相比差异极显著(P<0.01),但各干旱处理之间并无显著差异(P>0.05);在LS和MS处理下小紫珠ETo/RC与CK相比无显著差异(P>0.05),在SS处理下差异极显著(P<0.01),增加了20.31%;随着干旱胁迫程度的加剧,小紫珠DIo/RC显著增加,在LS、MS、SS处理下与CK相比分别增加了11.67%、37.29%、103.62%。
2.4 干旱胁迫对小紫珠叶绿素含量的影响
干旱胁迫对小紫珠叶绿素含量的影响如表4所示,随着干旱胁迫的加剧小紫珠叶绿素a逐渐下降,在MS和SS处理下与CK相比差异极显著(P<0.01),分别减少了20.59%、34.31%。小紫珠叶绿素b和叶绿素总量随着干旱胁迫加剧逐渐减少,与CK相比在SS处理下达到差异极显著(P<0.01),分别降低了28.21%、46.11%。小紫珠叶绿素a/b在MS和SS处理下与CK相比差异显著(P<0.05),分别减少了22.52%、36.25%。
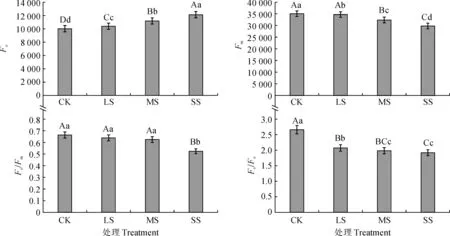
图3 干旱胁迫下小紫珠叶绿素荧光参数的变化Fig.3 The chlorophyll fluorescence parameters of the C. dichotoma under drought stress
处理 TreatmentφPoΨoφEoφDoCK0.722±0.003Dd0.558±0.007Cc0.416±0.001Dd0.278±0.005AaLS0.702±0.001Cc0.482±0.005Cc0.363±0.010Cc0.299±0.004AaMS0.690±0.002Bb0.446±0.007Bb0.311±0.002Bb0.349±0.014BbSS0.657±0.005Aa0.438±0.007Aa0.282±0.005Aa0.356±0.004Bc

表3 干旱胁迫下小紫珠PSⅡ比活性参数的变化
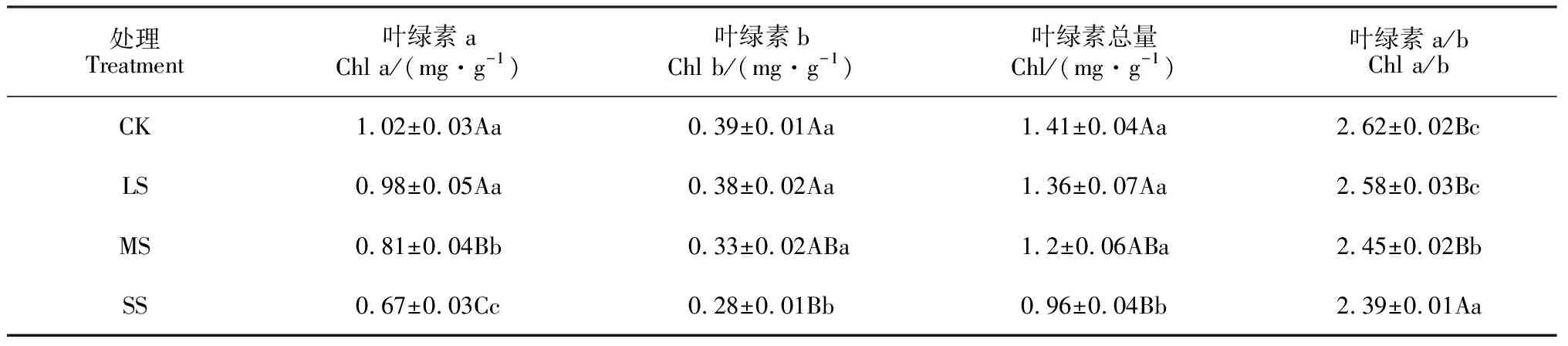
表4 干旱胁迫下小紫珠叶绿素含量及比值的变化
3 讨 论
植物光合作用受植物本身和环境因素的双重作用的影响,其中土壤水分是影响植物光合生理的重要因素[15]。植物气孔受土壤水分和根系信号的控制,干旱致使气孔关闭,限制了CO2的进入和水分的蒸发,导致植物光合作用的降低和蒸腾速率的下降[16]。前人研究表明,当植物处于逆境时,Pn、Ci和Gs同时降低时,主要是气孔限制引起的光合作用降低,如果Pn与Gs降低的同时伴随着Ci的升高,是由非气孔限制引起的光合作用降低[17]。试验结果表明,在轻度和中度干旱胁迫下,小紫珠的Gs明显下降,进入叶片的CO2减少,植物的光合受到抑制,说明此时光合作用的降低是由气孔限制引起的,在重度干旱胁迫下小紫珠的Gs和Pn持续下降,而Ci浓度升高,这是由于植物细胞受损产生的CO2增多,这说明在重度干旱胁迫下光合作用的下降是由光合机构活性降低的非气孔限制引起的。通过对小紫珠光合作用日进程研究发现,小紫珠在中度和重度干旱胁迫下出现光合“午休”现象,这是植物通过捕光色素复合体(LHCII)的可逆性脱离,导致光系统反应与光合效率下降及减少水分损失,使其在低水强光环境下避免受到损伤得以继续生存[18]。在轻度和中度干旱胁迫下,植物通过关闭气孔降低水分丧失来换取更大量的CO2同化,致使Tr降幅(18.95%、46.44%)大于Pn(12.46%、38.01%),导致小紫珠WUE下降幅度较小[19]。在重度干旱胁迫下植物水分失衡加剧,致使Tr降幅(52.76%)小于Pn(67.76%),导致小紫珠WUE显著下降,此结果与李志军等[20]研究一致。
光响应曲线反映了植物光合速率随光照强度增减变化的规律,方便了解植物光合能力及不同环境对其影响程度[21]。本试验应用直角双曲线修正模型对小紫珠光响应曲线拟合表明,在同一PAR下随着土壤含水量的降低,Pn降幅增大,说明干旱加重了小紫珠光抑制发生强度、降低了植物Rusbico活性及电子传递速率[22]。随着干旱胁迫的加剧小紫珠LSP和AQY显著降低,LCP显著增高,表明叶片利用弱光和强光的能力均降低[23]。这与倪霞等[24]在毛竹上的研究结果一致,说明在干旱胁迫下小紫珠把太阳能转化为净能量的能力降低。随着干旱胁迫加剧Rd显著降低,植物主要是通过降低呼吸作用对光合产物的过多消耗,以适应干旱的土壤条件。
叶绿素荧光技术是研究植物光合作用的无损伤探针,通过对快速叶绿素荧光参数的测定,分析其光系统Ⅱ的活性,可快速、准确地了解植物在不同环境条件下的生理状态[25]。试验结果发现,随着干旱胁迫的加剧小紫珠Fo逐渐升高,在中度和重度干旱胁迫下Fv/Fm和Fv/Fo显著降低,表明植物PSⅡ受到伤害,PSⅡ主动通过降低电子传递和减小光化学效率来响应CO2同化能力的降低,植物通过热耗散形式将过剩的光能释放以避免强光对光系统的破坏和损伤[26]。在试验中发现,随着干旱胁迫的加剧小紫珠φPo、Ψo和φEo逐渐降低,但φDo却明显上升,表明植物的自我保护机制启动,φEo明显的下降也表明了水分胁迫抑制了QA-的电子传递[27]。由于PSⅡ结构的破坏,电子传递和能量转化受到较大的影响,ABS/RC、TRo/RC、ETo/R、DIo/RC受到干旱胁迫后显著增加,这是由于单位面积的反应中心数目减少和剩余的有效反应中心效率增强,以更好地耗散电子传递链中的能量,说明反应中心吸收和捕获的光能除了电子传递外,大部分都以热量的形式耗散,这可能是小紫珠在干旱条件下的一种自我保护机制。
叶绿素的含量和比例在一定程度上能反映植物利用光能、累积生物量、适应和利用环境的能力。研究表明,干旱胁迫会抑制植物叶绿素的合成,并加速其分解,导致叶绿素含量的迅速下降[28]。试验发现,随着干旱胁迫的加剧,小紫珠叶绿素a、叶绿素b和叶绿素总量逐渐下降,土壤水分越低则降低越显著,表明轻度和中度干旱胁迫对小紫珠光合器官活性与功能的影响较为轻微,在重度干旱胁迫下则影响较大,植物的光合器官活性和功能明显受到伤害,这与光合参数的研究结果相一致。随着干旱胁迫的加剧,小紫珠叶绿素a/b逐渐下降,在中度和重度干旱胁迫下达到差异极显著(P<0.01),说明叶绿素a的降幅要大于叶绿素b,干旱并没有显著影响植物的叶绿素b含量,叶绿素总量的降低主要是由于叶绿色a的下降引起的,表明了干旱下小紫珠的光系统反应中心比捕光天线系统更易受损,这可能与膜脂过氧化程度加大有关[29]。
综合上述分析可知,在干旱胁迫下,小紫株的Pn、Gs以及光化学转化效率和CO2同化能力的相关参数均受到不同程度的抑制,说明各种干旱胁迫下光合速率的降低既有气孔因素也有非气孔因素,但在轻度和中度干旱胁迫下光合速率降低的主要原因为气孔因素,而在重度干旱胁迫下其主要原因为非气孔因素。
参考文献:
[1] 王 缓, 盛连喜, 孙弘颜.中国水资源现状分析与可持续发展对策研究[J].水资源与水工程学报, 2008,19(3): 10-15.
WANG Y, SHENG L X, SUN H Y,etal. Analysis of present situation of water resources and countermeasures for sustainble development in China[J].JournalofWaterResources&WaterEngineering, 2008,19(3): 10-15.
[2] 金英杰.浅谈中国水资源短缺的现状及节水措施[J].中国高新技术企业, 2009, (15): 7-39.
[3] 张建国, 李吉跃, 沈国舫.树木耐早特性及其机理研究[M].北京:中国林业出版社, 2000.
[4] 李春燕, 王进鑫, 薛 设.干旱及复水对侧柏幼苗叶片抗氧化酶活性的影响[J].西北林学院学报2015,30(2): 33-37.
LI C Y, WANG J X, XUE S. Effect of drought and rewatering on the activity of protect enzymes in the leaves ofPlatycladusorientalisseedings[J].JournalofNorthwestForestryUniversity, 2015,30(2): 33-37.
[5] 柴胜丰, 唐健民, 王满莲, 等.干旱胁迫对金花茶幼苗光合生理特性的影响[J].西北植物学报, 2015,35(2): 322-328.
CHAI S F, TANU J M, WAND M L,etal. Photosynthetic and physiological characteristics ofCamelliapetelotiiseedlings under drought stress[J].ActaBotanicaBoreali-OccidentaliaSinica, 2015,35(2): 322-328.
[6] 郭有燕, 刘宏军, 孔东升, 等.干旱胁迫对黑果枸杞幼苗光合特性的影响[J].西北植物学报, 2016,36(1): 124-130.
GUO Y Y, LIU H J, KONG D S,etal. Effect of drought stress on photosynthesis characteristics oflyciumruthenicumseedlings[J].ActaBotanicaBoreali-OccidentaliaSinica, 2016,36(1): 124-130.
[7] 谷衍川, 丰震, 李承水, 等.小紫珠种群的表型遗传多样性研究[J].山东林业科技, 2013, (2): 5-9.
GU Y C, FENG Z, LI C S,etal. Resaerch in variation characteristics of phenotypic traits in the SpeciesCallicarpadichotoma(Lour.)K. Koch[J].JournalofShandongForestryScienceandTechnology, 2013, (2): 5-9.
[8] 房丽琴.大叶紫珠和小紫珠的扦插繁殖技术[J].安徽农业科学, 2010,38(24): 13 175-13 176.
FANG L Q. Research on cutting propagation ofCallicarpamacropyllaandCallicarpadichouma[J].JournalofAnhuiAgriculturalSciences, 2010,38(24): 13 175-13 176.
[9] 罗桑卓玛, 辛福梅, 杨小林, 等.干旱胁迫对香柏幼苗生长和生理指标的影响[J].西北农林科技大学学报(自然科学版), 2015,43(5): 51-70.
LUOSANG Z M, XIN F M, YANG X L,etal. Effect of drought stress on growth and physiological indicators ofSabinapingiivar.wilsoniiseedlings[J].JournalofNorthwestA&FUnivcrsity(Nat.Sci.Ed.), 2015,43(5): 51-70.
[10] 王建华, 任士福, 史宝胜, 等.遮荫对连翘光合特性和叶绿素荧光参数的影响[J].生态学报, 2011,31(7): 1 811-1 817.
WANG J H, REN S F, SHI B S,etal. Effects of shades on the photosynthetic characteristics and chlorophyll fluorescence parameters ofForsythiasuspensa[J].ActaEcologicaSinica, 2011,31(7): 1 811-1 817.
[11] 胡 义, 胡庭兴, 胡红玲, 等.干旱胁迫对香樟幼树生长及光合特性的影响[J].应用与环境生物学报, 2014,20(4): 675-682.
HU Y, HU T X, HU H L,etal. Effects of drought stress on growth and photosynthetic characteristics ofCinnamomumcamphorasaplings[J].ChineseJournalofAppliedEcology, 2014,20(4): 675-682.
[12] 叶子飘.光合作用对光合CO2响应模型的研究进展[J].植物生态学报, 2010,34(6): 727-740.
YE Z P. A review on modeling of responses of photosynthesis to light and CO2[J].ChineseJournalofPlantEcology, 2010,34(6): 727-740.
[13] 魏晓东, 陈国祥, 施大伟, 等.干旱胁迫对银杏叶片光合系统Ⅱ荧光特性的影响[J].生态学报, 2012,32(23): 7 492-7 500.
WEI X D, CH G X, SHI D W,etal. Effects of drought on fluorescence characteristics of photosystem Ⅱ in leaves ofGinkgobiloba[J].ActaEcologicaSinica, 2012,32(23): 7 492-7 500.
[14] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社, 2000.
[15] SOFO A, DICHIO B, MONTANARO G,etal. Photosynthetic performance and light respose of two olive cultivars under different water and light regimes[J].Photosynthetica, 2009,47(4): 602-608.
[16] DAVIES W J, ZHANG J. Root signals and the regulation of growth and development of plants in drying soil[J].AnnualReviewofPlantPhysiologyandPlantMolecularBiology, 1991,42(1): 55-76.
[17] FARQUHAR G D, SHARKEY T D. Stomatal conductance and photosynthesis[J].AnnualReviewofPlantPhysiology, 1982,33(1): 317-345.
[18] CHEN Y, XU D Q. Two patterns of leaf photosynthetic response to irradiance transition from saturating to limiting one in some plant species[J].NewPhytologist, 2006,169(4): 789-798.
[19] 姬兰柱, 肖冬梅, 王 森.模拟水分胁迫对水曲柳光合速率及水分利用效率的影响关[J].应用生态学报, 2005,16(3): 1 516-1 524.
JI L Z, XIAO D M, WANG S. Effects of simulated water stress on photosynthesis rate and WUE ofFraxinusmandshurica.[J].ChineseJournalofAppliedEcology, 2005,16(3): 1 516-1 524.
[20] 李志军, 罗青红, 伍维模, 等.干旱胁迫对胡杨和灰叶胡杨光合作用及叶绿素荧光特性的影响[J].干旱区研究, 2009,26(1): 45-52.
LI Z J, LUO Q H, WU W M,etal. The Effects of droughtstress on photosynthetic and chlorophyll fluorescence characteristics ofPopuluseuphraticaand p.pruinosa[J].AridZoneResearch, 2009,26(1): 45-52.
[21] 叶子飘, 于 强.光合作用光响应模型的比较[J].植物生态学报, 2008,32(6): 1 356-1 361.
YE Z P, YU Q. Comparison of new and several classical models of photosynthesis in response to irradiance[J].JournalofPlantEcology, 2008,32(6): 1 356-1 361.
[22] 陆燕元, 马焕成, 李昊民, 等.土壤干旱对转基因甘薯光合曲线的响应[J].生态学报, 2015,35(7): 2 155-2 160.
LU Y Y, MA H C, LI H M,etal. Light response characteristics of photosynthetic of transgenic sweet potato under drought stress[J].ActaEcologicaSinica, 2015,35(7): 2 155-2 160.
[23] XIA J B, ZHANG S Y, ZHANG G C,etal. Critical responses of photosynthetic efficiency inCampsisradicans(L.) Seem to soil water and light intensities[J].AfricanJournalofBiotechnology, 2011,10(77): 17 748-17 754
[24] 倪 霞, 曹永慧, 周本智, 等.干旱处理对毛竹光响应的影响:基于4种模型比较分析[J].林业科学研究, 2017,30(3): 465-471.
NI X, CAO Y H, ZHOU B Z,etal. Light response ofPhyllostachysedulisunder drought stress: based on 4 models[J].ForestResearch, 2017,30(3): 465-471.
[25] JIANG C D, GAO H Y, ZOU Q. Changes of donor and accepter side in photosystem 2 complex induced by iron deficiency in attached soybean and maize leaves[J].Photosvnthetia, 2003, 41: 267-271.
[26] 卜令铎, 张仁和, 常 宇, 等.苗期玉米叶片光合特性对水分胁迫的响应[J].生态学报, 2010,30(5): 1 184-1 191.
BU L Z, ZHANG R H, CHANG Y,etal. Response of photosynthetic characteristics to water stress of majze leaf in seeding[J].ActaEcologicaSinica, 2010,30(5): 1 184-1 191.
[27] 冯胜利, 马富裕, 方志刚, 等.水分胁迫对加工番茄光系统Ⅱ的影响[J].干旱地区农业研究, 2009,27(1): 163-167.
FENG S L, MA F Y, FANG Z G,etal. Effects of water stress on photosystem Ⅱ in tomato[J].AgriculturalResearchintheAridAreas, 2009,27(1): 163-167.
[28] 陈卫元, 曹 晶, 姜卫兵.干旱胁迫对红叶石楠叶片光合生理特性的影响[J].中国农学通报, 2007,23(8): 217-220.
CHEN W Y, CAO J, JIANG W B. Effects of drought stress on photosynthetic characteristics ofPhotiniafraserydress[J].ChineseAgriculturalScienceBulletin, 2007,23(8): 217-220.
[29] 王海珍, 韩 路, 徐雅丽, 等.土壤水分梯度对灰胡杨光合作用与抗逆性的影响[J].生态学报, 2017,37(2): 433-442.
WANG H Z, HAN L, XU Y L,etal. Effects of soil water gradient resistance ofPopuluspruinosaon photosynthetic characteristics and stress in the Tarim Basin,China[J].ActaEcologicaSinica, 2017,37(2): 433-442.
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28
阅读(科学探秘)(2020年8期)2020-11-06
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
中国果业信息(2019年1期)2019-01-05
生物学教学(2017年9期)2017-08-20
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
中国铸造装备与技术(2015年5期)2015-12-10
少儿科学周刊·儿童版(2015年10期)2015-11-07
