
乌金猪mtDNA D-Loop区的遗传多样性*
2018-05-17 08:27:56王志敏徐亚欧徐珑洋王英明黄卫平
吉首大学学报(自然科学版) 2018年2期
王志敏,徐亚欧,孙 艳,杨 磊,徐珑洋,王英明,黄卫平
(1.西南民族大学生命科学与技术学院,四川 成都 610041;2.凉山州畜牧兽医科学研究所,四川 凉山州 615042)
乌金猪原产于云贵高原,是我国优良的地方遗传资源之一,主要分布在云南、贵州和四川交界的大部分地区.毛色主要有黑色和棕黄色,因产地的不同在各地的叫法也不同.在四川的凉山彝族自治州被称作凉山猪,主要分布于凉山州的西昌、昭觉和美姑等地.贵州的柯乐猪、威宁猪,云南的大河猪等均属于乌金猪不同类群.1976年云南、贵州和四川联合调查认为,上述不同名称的猪种属同种异名,将其统称为乌金猪.乌金猪具有体型健壮、后驱发达、能适应高寒气候和粗放饲养,适宜高原牧场繁衍,形成了肉质鲜嫩,口感好,在云南、贵州和四川的冕宁等地有用做火腿的习惯[1-3].
对乌金猪的起源及不同类群的遗传关系研究多基于其外貌特征,分布地域,群体变化,生产性能,历史考证等方法进行.利用现代分子生物学方法对乌金猪不同类群间的遗传亲缘关系研究还未见报道.动物mtDNA具有广泛的种内和种间多态性,且为母性遗传,在亲缘关系相近的物种间其进化速度比核基因快,因而它为种群遗传学和进化遗传学在分子水平上的研究提供了理想的研究对象.并已广泛的应用于动物遗传学、遗传分化、种群遗传结构分析、原种鉴定、分子生态学等方面[4-7].本研究利用mtDNA D-loop的多态性对分布于与云南(大河猪)、贵州(柯乐猪)和四川(凉山猪)乌金猪的不同类群的亲缘关系进行比较研究,为今后乌金猪不同类群遗传资源的保护与开发利用提供理论依据.
1 材料与方法
1.1 实验材料
实验用乌金猪血液样本由四川省凉山彝族自治州畜牧科学研究院提供.选取健康成年乌金猪(不区分公母)的3个类群,四川凉山猪27只(昭觉11只,美姑13只,西昌3只),贵州毕节地区柯乐猪21只,云南富源县大河猪20只.用内含EDTA2K抗凝剂的试管自颈静脉采血,血液样本低温保存带回实验室备用.
1.2 主要试剂
2*Tag酶PCR Master Mix、DNase/RNase-free去离子水、D2000 DNA Marker、天根基因组DNA提取试盒和琼脂糖Agarose,均购自成都康迪生物技术有限公司.
1.3 基因组DNA的提取及检测
利用天根生物试剂有限公司离心柱型血液基因组DNA提取试剂盒,从EDTA抗凝全血中提取基因组DNA.紫外分光光度计检测DNA样品D 260/280值及浓度.配制1.5%琼脂糖平板凝胶,电泳缓冲液为0.5×TBE,溴化乙锭染色,电压为120 V,电泳时间为30 min,凝胶成像系统观察、保存.
1.4 引物的设计与合成
根据猪线粒体DNA序列(AF034253)[8],用Primer Premier5.0软件设计扩增引物1,扩增片段包括完整的D-loop,引物序列见表1,交由成都擎科梓熙生物有限公司合成.根据成都擎科梓熙生物有限公司返回的测序结果运用Primer Premier5.0设计引物2对突变频率较高的区域进行扩增,然后将PCR产物送往公司进行测序,设计引物见表1.
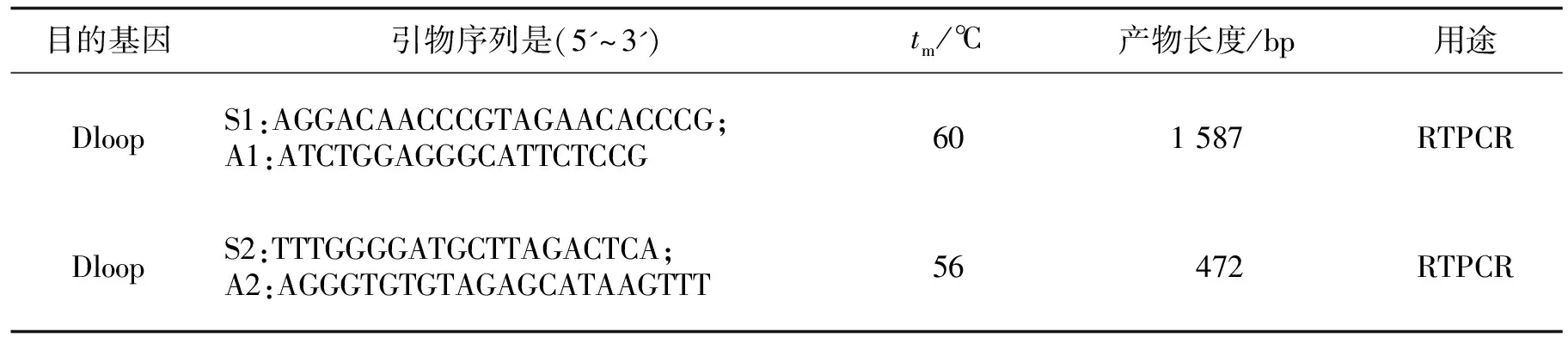
表1 引物信息Table 1 Primer Information
PCR扩增体系共50 μL:Taq聚合酶25 μL,ddH2O19 μL,cDNA2 μL,上下游引物各2 μL.PCR反应程序:94 ℃预变性(4 min);94 ℃变性(20 s),Tm退火(60 s),72 ℃延伸(90 s),共39个循环,72 ℃延伸(10 min).取5 μLPCR产物,用1.5%琼脂糖凝胶进行检测,将产物送往擎科进行测序(1 587 bp产物进行双向测序,472 bp产物进行正向测序).
1.5 数据处理与分析
用vectorNTI软件对测定的序列进行比对后截取突变频率集中的序列,DNAStar软件统计截取序列的长度和碱基组成,MEGA Ver5.0 软件计算类群间的 Kimura 双参数距离[9]并以邻接法(Neighbor-Joining,NJ) 构建类群间的进化树;并进行多态位点、核苷酸多样性分析.用clustalw对序列聚类,用TreeView软件制作网络关系图.
2 结果与讨论
2.1 mtDNA D-Loop区及其突变位点富集区的扩增
5个地区乌金猪类群完整mtDNA D-loop区及其突变位点富集片段的PCR扩增序列通过1.5%琼脂糖凝胶电泳检测后,与预期大小相符,见图1及图2.
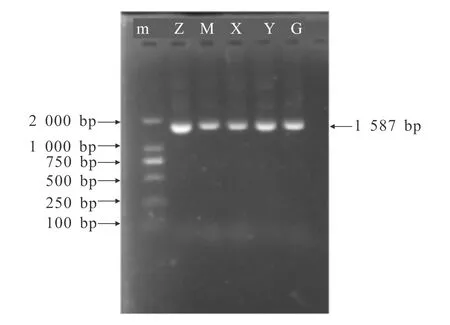
m— DNA Marker;Z—昭觉乌金猪;M—美姑乌金猪;X—西昌乌金猪;Y—云南乌金猪;G—贵州乌金猪.图1 5个类群乌金猪mtDNA D-Loop区PCR扩增结果Fig.1 PCR Amplification Result of mtDNA D-Loop of 5 Wujin Pigs
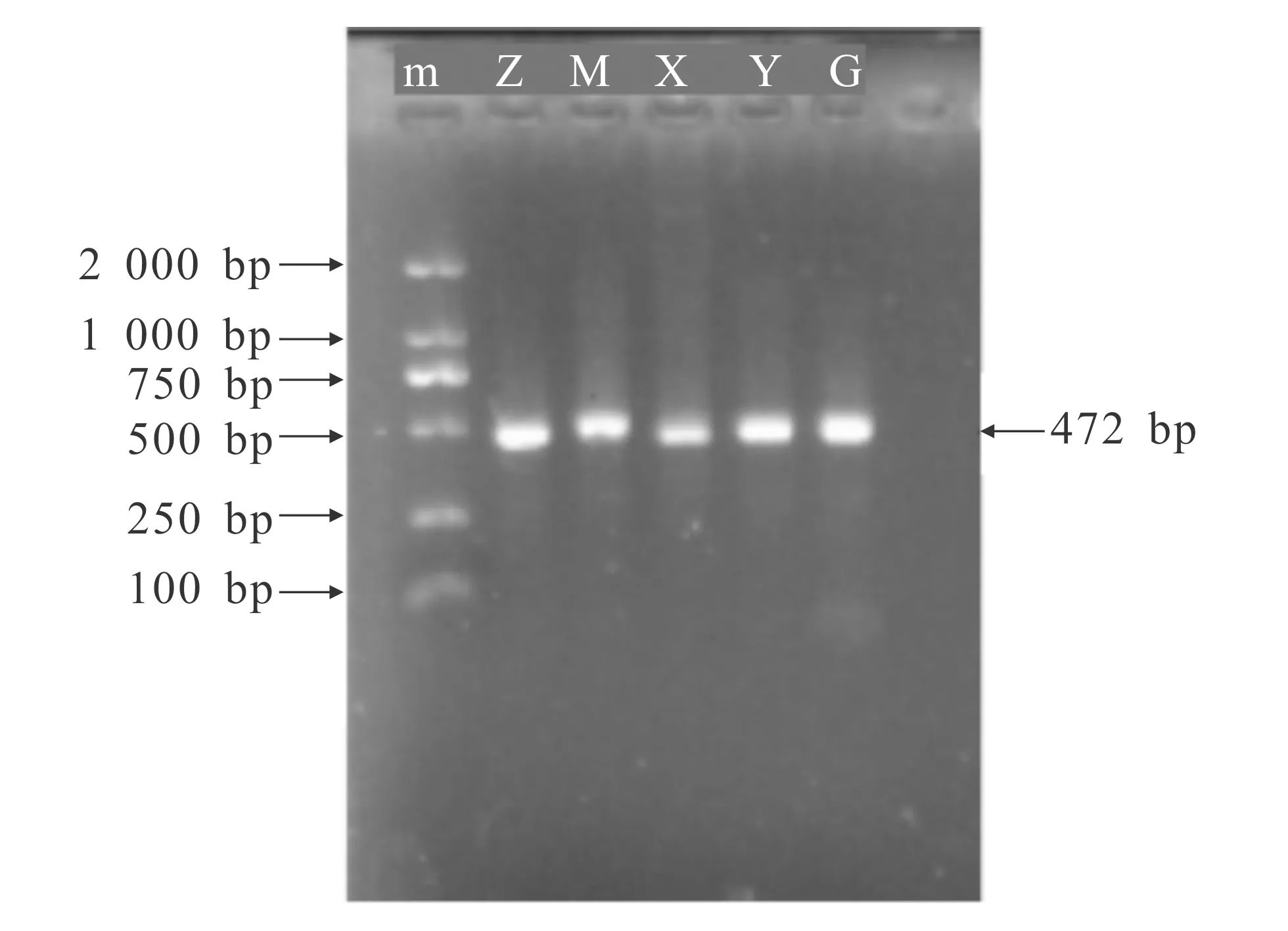
m—DNA Marker;Z—昭觉乌金猪;M—美姑乌金猪;X—西昌乌金猪;Y—云南乌金猪;G—贵州乌金猪.图2 5个类群乌金猪mtDNA D-Loop区突变富集区PCR扩增结果Fig.2 PCR Amplification Results of mtDNA D-Loop Region Mutation Enrichment Region of 5 Wujin Pigs
2.2 序列的长度与碱基的组成
PCR扩增产物送往擎科进行测序.用vectorNTI软件对测定的序列进行比对后截取同源序列,用DNAStar软件统计截取序列的长度为249 bp,5个地区乌金猪的A,G,T,C碱基含量见表2.
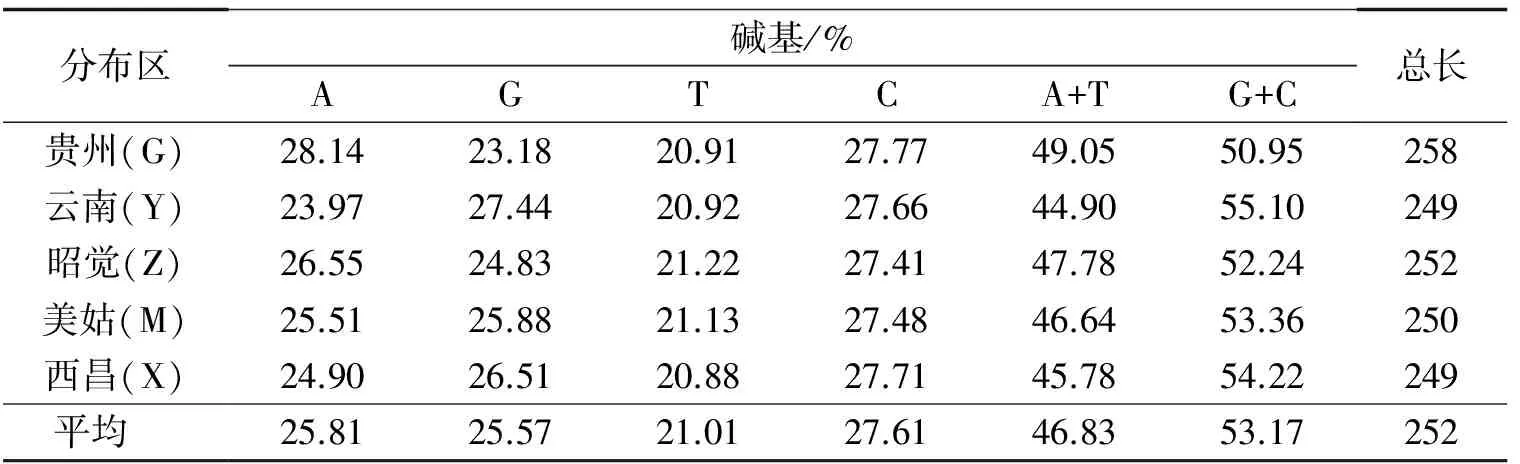
表2 5个类群乌金猪mtDNA D-Loop区突变富集区的碱基频率Table 2 Base Frequency of the mtDNA D-Loop Region of 5 Wujin Pigs
由表2可知,A,G,T,C碱基的平均为25.81%,25.57%,21.01%和27.61%,且G+C的平均(53.17%)高于A+T的平均(46.83%),这表明该区域结构比较稳定.
2.3 核苷酸序列多态性
在测序的乌金猪mtDNA D-loop区全序列共46个突变位点,简约信息位点有5个,单一多态位点37个,其中以A/G突变为主.就变异位点而言,贵州及四川类群的变异位点数较多,而云南的变异较小,其结果见表3.
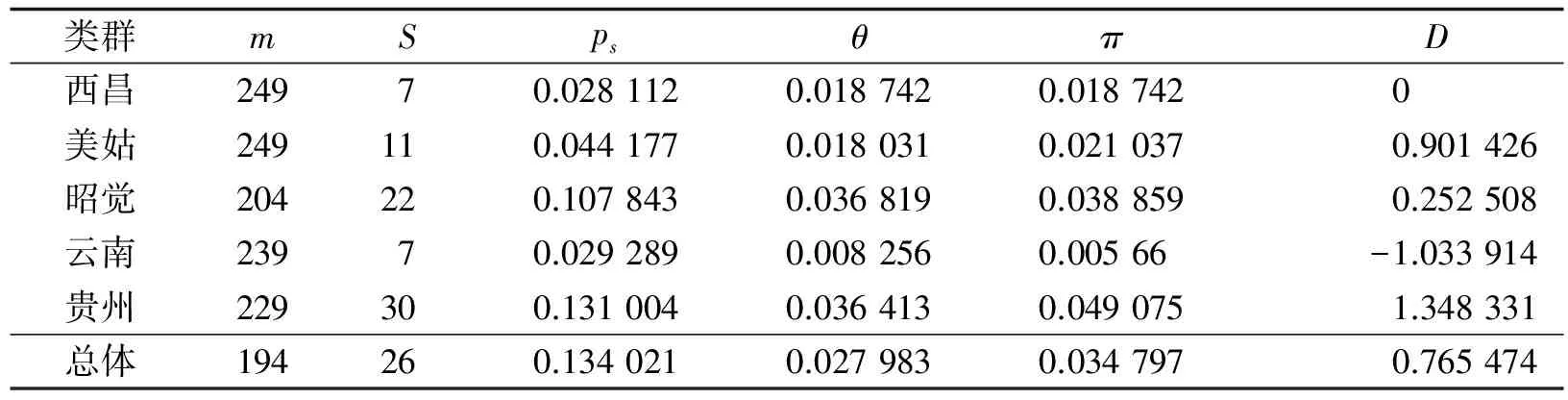
表3 核苷酸序列多态性分布情况Table 3 Nucleotide Sequence Polymorphism Distribution
注:m为分析位点数;S为变异位点数;ps=s/m;θ=ps/a1;π为核苷酸多态性;D为Tajima检验统计量.
2.4 乌金猪种群间的遗传距离和进化树的构建
用MEGA5.0软件基于Kimura 双参数计算乌金猪5个类群间的遗传距离见表4,并用NJ法构建进化树见图3.
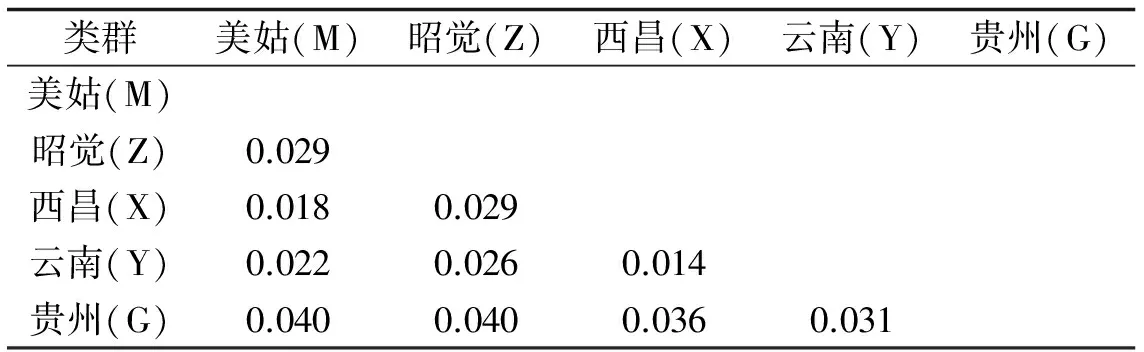
表4 5个类群乌金猪的种间的平均遗传距离Table 4 Average Genetic Distance of 5 Wujin Pigs
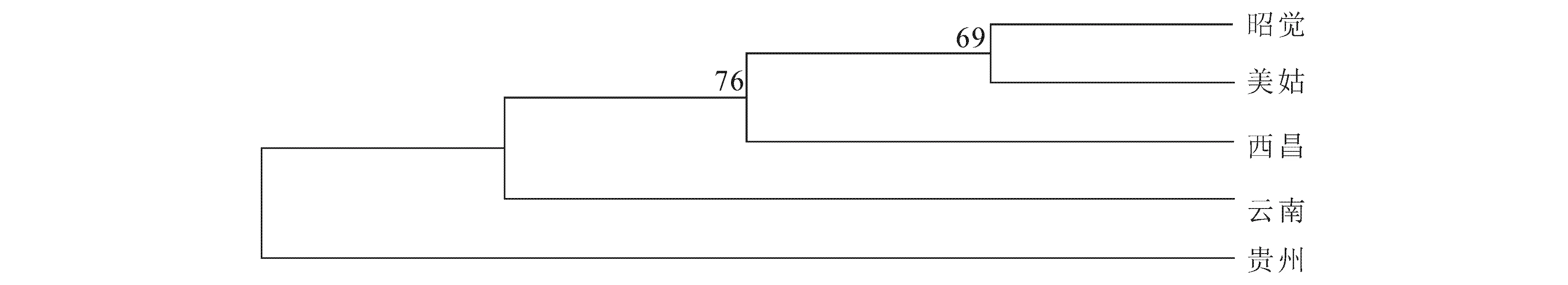
图3 基于Kimura 双参数遗传距离的乌金猪类群间的NJ进化树Fig.3 NJ Phylogenetic Tree Based on Kimura's Two-Parameter Genetic Distance Among Wujin Pigs
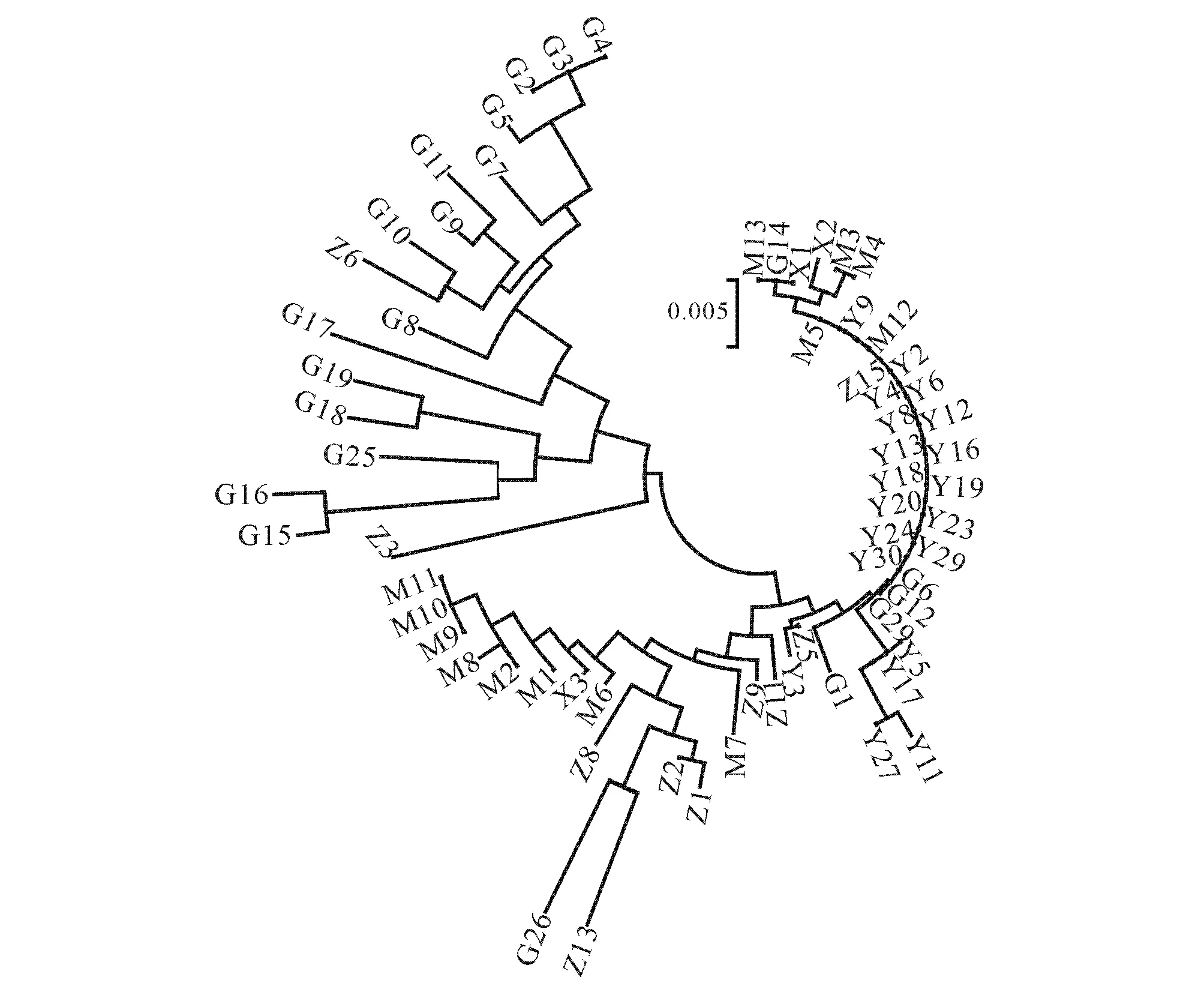
图4 5个地区乌金猪网络关系进化Fig.4 Evolution of Wujin Pig Network in Five Areas
表4中贵州乌金猪(柯乐猪)与四川乌金猪(凉山猪)的美姑、昭觉、西昌类群的遗传距离较大,再由图3也可看出柯乐猪较四川凉山猪类群距离较远.云南乌金猪(大河猪)与四川乌金猪(凉山猪)的美姑、昭觉、西昌类群的遗传距离相对较小,由图3可看出大河猪与四川乌金猪(凉山猪)的美姑、昭觉、西昌类群遗传距离相对较近.图3显示凉山猪的3个类群(昭觉、美姑、西昌)首先聚为一类,然后四川乌金猪(凉山猪)与云南乌金猪(大河猪)聚为一类,四川乌金猪(凉山猪)与贵州乌金猪(柯乐猪)最后聚类,亲缘关系最远.
根据5个地区乌金猪的序列,用Clustalx进行聚类,再用TreeView制作5个地区乌金猪网络关系进化图(图4).由图4可见所研究的3个乌金猪类群可以分为2个大类,贵州乌金猪(柯乐猪)自成一类,云南乌金猪(大河猪)与四川乌金猪(凉山猪)为一个类群,说明3个地区的乌金猪可能有2个母系起源.且在2个母系起源的类群中,相互之间也有血缘的交叉.
3 结论与分析
本研究对分布于西南地区云南、贵州和四川的地方猪种乌金猪的mtDNA D-loop区进行PCR扩增,经测序发现突变位点主要集中与290 bp的区段,分析该片段的碱基组成,发现A+T平均低于G+C平均,说明该区段结构比稳定.并对该序列进行核苷酸多态性统计,全序列共46个突变位点,简约信息位点有5个,单一多态位点37个,其中以A/G突变为主.基于Kimura双参数计算类群间的遗传距离并构建NJ进化树,由进化树可见四川乌金猪(凉山猪)的3个类群(昭觉、美姑、西昌)首先聚为一类,然后四川乌金猪(凉山猪)与云南乌金猪(大河猪)聚为一类,最后四川乌金猪(凉山猪)与贵州乌金猪(柯乐猪)聚类.赵思思等[10]对西南型地方猪品种资源的变化的研究表明,1986年出版的《中国猪品种志》中乌金猪主要包含柯乐猪、威宁猪、大河猪和凉山猪类群,2011年出版的《中国畜禽遗传资源志——猪志》中,柯乐猪、大河猪、昭通猪和凉山猪统称为乌金猪.其对乌金猪的类群做了一定的调整,从20世纪80年代到21世纪初乌金猪的数量增加了1倍以上.由本研究的结果显示,3个乌金猪类群可以分为2个母系起源,贵州乌金猪(柯乐猪)自成一类,云南乌金猪(大河猪)与四川乌金猪(凉山猪)为一个类群,这就从分子水平证实了大河猪、凉山猪属同种异名的2个类群.
郭宏宇等[11]对贵州省的关岭猪、柯乐猪和糯谷猪的研究表明,其中柯乐猪及糯谷猪均属于乌金猪的地方类群.兰宏等[12]对西南地区家猪和野猪mtDNA遗传多样性研究表明,西南地区的猪起源于一个共同的祖先,在品种形成的早期可能受到创立者效应的制约.如若西南地区猪起源于一个共同祖先,那么乌金猪在贵州、四川和云南等地的遗传关系就显而易见,总体而言具有较近的亲缘关系,但由于各地群众的生产生活方式的不同,经过长期的选育由形成了各自的特点,从而满足当地人们的生活需要.本研究结果对于乌金猪的保种与开发,尤其是对四川乌金猪(凉山猪)的保护与开发提供了理论基础,也为四川、云南和贵州各地乌金猪的不同类群的合理利用提供了理论依据.
参考文献:
[1] 黄启昆,王玉嵩,王守信,等.云南省家畜家禽品种志[M].昆明:云南科技出版社,1987:18-23.
[2] 郭海涛,郭成裕,李军鹏,等.我国地方猪品种——乌金猪的生理特性研究[J].现代畜牧兽医,2008(10):17-18.
[3] 国家畜禽遗传资源委员会.中国畜禽遗传资源志(猪志)[M].北京:中国农业出版社,2011.
[4] 杨明生,熊邦喜.动物MTDNA的研究及在鱼类生态学中的应用[J].孝感学院学报,2005(3):23-27.
[5] 李洪涛,顾为望,吴清洪,等.实验用藏猪和巴马小型猪线粒体DNA控制区碱基序列比较[J].郑州大学学报(医学版),2008,43(5):933-935.
[6] 汪泰初,刘朝良,肖林珍.线粒体基因组(MTDNA)的研究进展[J].安徽农业科学,2006,34(10):2 068-2 068.
[7] 潘宝平,卜文俊.线粒体基因组的遗传与进化研究进展[J].生物学通报,2005,40(8):1-3.
[8] 赵兴波,李 宁.猪线粒体DNA D-loop PCR-RFLP分析[J].中国农业大学学报,1998(5):13-17.
[9] 李彬彬,黄培春,钟复光.生物学软件在线粒体DNA序列多态性分析中的应用[J].生物信息学,2010,8(2):153-155.
[10] 赵思思,贾 青,胡慧艳,等.西南型地方猪品种资源的变化[J].贵州农业科学,2016,44(10):91-94.
[11] 郭宏宇,林家栋.利用微卫星DNA标记分析贵州3个地方猪种的遗传多样性[J].安徽农业科学,2009,37(8):77-80.
[12] 兰 宏,王 文,施立明.西南地区家猪和野猪MTDNA遗传多样性研究[J].Journal of Geneticss & Sgenomics,1995(1):28-33.
猜你喜欢
VOGUE服饰与美容(2022年5期)2022-05-01 10:38:53
小猕猴学习画刊·下半月(2022年2期)2022-04-16 16:06:33
——探访煤炭博物馆
奇妙博物馆(2022年3期)2022-03-23 05:16:20
小学生(看图说画)(2019年9期)2019-09-09 02:17:02
江苏农业科学(2019年5期)2019-09-02 14:01:46
中成药(2018年12期)2018-12-29 12:25:34
环球时报(2018-12-14)2018-12-14 08:10:48
闽南风(2018年9期)2018-10-31 14:48:46
广东农业科学(2017年5期)2017-08-29 10:37:31
应用海洋学学报(2014年1期)2014-11-22 07:17:44
