
内质网应激对大鼠局灶性脑缺血再灌注后听皮层损伤的作用*
2018-05-10 11:46:33时美娟宋永周
重庆医学 2018年11期
吕 哲,张 颖,时美娟,孟 晴,宋永周,路 虹△
(河北医科大学第二医院:1.耳鼻咽喉科;2.骨科,石家庄 050000)
研究表明脑缺血再灌注时缺血缺氧、自由基蓄积及三磷酸腺苷耗竭等都可能导致内质网应激(endoplasmic reticulum stress,ERS)[1-2]。过度ERS可以引起神经细胞凋亡,进一步加重脑损伤。据报道听力缺失与脑卒中密切相关[3]。众所周知,听皮层是听觉系统的最高级中枢。初级神经元的退行性变及凋亡将导致中枢对传入信息的感知能力下降,对声音信息的认知和处理的能力下降。以往对缺血再灌注损伤导致的听力损伤研究多集中于外周听觉系统,而对中枢病变研究较少[4-6]。本研究通过建立局灶性脑缺血再灌注模型,观察内质网应激相关因子葡萄糖调节蛋白78(glucose regulated protein 78,GRP78)、天冬氨酸特异性半胱氨酸蛋白酶-12(Caspase-12)在听皮层的表达,阐述脑缺血再灌注对听力损害的可能机制。
1 材料与方法
1.1实验动物 健康雄性清洁级SD大鼠30只,体质量250~280 g,无耳毒性药物使用史,无噪声暴露史,耳廓反射灵敏,由河北医科大学实验动物中心提供。动物许可证号:SCXK(冀)2013-1-003。饲养条件:以标准饲料喂养,恒温20~25 ℃,昼夜交替。采用随机数字表法把实验动物分为两组:缺血再灌注组(I/R组)15只,假手术组(Sham组)15只。
1.2仪器与试剂 ICS-CHARTR-EP型ABR电位仪(麦德森医疗器),Tunel试剂盒(Roche公司),三苯氯化四氮唑(TTC, Sigma公司),兔抗鼠Caspase-12抗体、GRP78抗体(Santa Cruz公司),免疫组织化学试剂盒(渤海生物工程有限公司),羊抗兔IgG荧光抗体(碧云天生物技术有限公司)。
1.3方法
1.3.1MACO模型建立 大鼠以10%水合氯醛全身麻醉后,依次分离颈总动脉和颈内、颈外动脉,结扎颈外动脉远端,颈外动脉上剪一小口,把标记好的栓线从切口处插入颈内动脉,随后向上深入,大约距颈总动脉分叉处(18.5±0.5)mm,此时堵住大脑中动脉开口,随即结扎固定,计时60 min,然后轻轻取出线栓。Sham组只进行血管分离并不插入线栓。24 h后进行检测和处死、取材。
1.3.2神经功能缺失评分 采用改良Longa法,即单盲 6级5分法进行评分。0分:无神经损伤症状表现;1分:提尾时不能伸展对侧的前肢;2分:提尾时对侧的前肢屈曲;3分:行走时轻度向对侧转圈;4分:行走时向对侧严重转圈;5分:向对侧跌倒,不能自主行走。
1.3.3听性脑干反应(auditory brainstem response,ABR)测试 分别对造模前和造模24 h后的两组动物进行检测。检测方法为:水合氯醛麻醉后置于隔声屏蔽室内,选用听性脑干诱发电位检测仪,采用短声为刺激声,增益为50 k,速率为每秒21.1次,极性为交替波,观察时程10 ms[7]。将初始刺激强度定为90 dB SPL,随后以5 dB SPL逐渐递减。直到递减至出现2次ABR Ⅲ波的波形不一致,此时增加5 dB SPL进行检测,当重复波形出现一致时,即可记为ABR的反应阈值。
1.3.4脑梗死体积测定 麻醉后每组取5只大鼠断头取脑,将其均匀切成5片冠状切片,随即浸入2%TTC溶液中染色30 min,在多聚甲醛溶液中固定24 h,取出拍照,计算梗死体积。脑梗死体积百分比=[总梗死体积-(梗死侧半球体积-梗死对侧半球体积)]/梗死对侧半球体积×100%。
1.3.5HE染色观察听皮层组织学改变 大鼠全身麻醉后暴露心脏,自心尖处将灌注针插入,以4%多聚甲醛灌注,固定后取脑。准确取材听皮层区域(参照《大鼠脑立体定位图谱》),组织置于固定液中24 h以上,之后石蜡包埋、切片。切片常规经脱蜡、初染、复染、脱水、封片等工序处理后进行观察。
1.3.6原位缺口末端标记(TUNEL)法检测凋亡神经细胞 常规处理制备好的石蜡切片后,按试剂盒说明书进行操作后,置于光镜下观察,若细胞核内为棕黄色即为阳性细胞。随机选择并计算5个高倍视野中的凋亡指数(AI)。AI=(凋亡细胞数/细胞总数)×100%。
1.3.7免疫组织化学观察听皮层GRP78、Caspase-12表达 将经抗原修复,灭活酶后的石蜡切片,滴加一抗、二抗,DAB显色及复染,封固后于显微镜下观察。随机选取5个不同的视野进行图像分析(采用Image-Pro Plus6.0软件)。
1.3.8蛋白免疫印迹(Western blot)法检测GRP78、Caspase-12蛋白表达 麻醉后取出大鼠梗死侧听皮层脑组织,迅速制备脑组织匀浆,在离心机中离心30 min后取上清液,进行蛋白定量。按说明书操作变性、上样之后,电泳、转膜,将其置入5%脱脂奶粉室温封闭1 h,分别加入兔抗Caspase-12多克隆抗体,兔抗GRP78多克隆抗体,孵育过夜后洗膜,再加入羊抗兔IgG荧光抗体,孵育1 h,应用远红外荧光扫描成像系统,以β-actin的表达为内参照,测定目标带单位密度, GRP78/β-actin、Caspase-12/β-actin比值代表蛋白相对表达水平。

2 结 果
2.1神经功能缺失评分、脑梗死体积比较 I/R组表现出不同程度的脑缺血性损伤,Sham组无行为学改变,经TTC染色后Sham组脑组织显示出均匀的红色。与Sham组比较,I/R组脑组织显示为大范围苍白色梗死区域,两组脑梗死体积、神经功能缺失评分、AI比较差异均具有统计学意义(P<0.01),见表1。
2.2ABR反应阈 造模前两组ABR反应阈比较差异无统计学意义(P>0.05),造模后两组比较差异有统计学意义(P<0.01)。I/R组造模前后差异有统计学意义,Sham组造模前后差异无统计学意义(P>0.05),见表2。
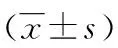
表1 大鼠神经功能缺损评分、脑梗死体积、AI比较

表2 ABR检测结果
2.3听皮层HE染色检测组织学改变 经HE染色光学显微镜下观察发现,Sham组神经元边界清楚,结构完整,形态正常,高倍镜下核仁清晰,细胞核圆而大, I/R组正常组织结构消失,细胞排列紊乱,细胞周围空泡变,细胞质嗜伊红,细胞核固缩、深染(图1)。
2.4TUNEL法检测凋亡神经细胞 缺血再灌注后24 h后,I/R组阳性细胞明显增多,细胞形态上有改变,两组AI比较差异具有统计学意义。提示缺血再灌注可以诱导细胞凋亡(表1、图2)。

A:Sham组;B:I/R组
图1听皮层神经细胞HE染色(×200)

A:Sham组;B:I/R组;C:两组AI比较,*:P<0.01,与Sham组比较
图2听皮层神经细胞TUNEL染色(×200)

A、D:Sham组;B、E:I/R组;C、F:两组GRP78、Caspase-12蛋白表达比较;*:P<0.01,与Sham组比较
图3听皮层神经细胞GRP78 、Caspase-12免疫组织化学染色结果(×200)
2.5免疫组织化学观察听皮层GRP78、Caspase-12表达 GRP78抗原阳性细胞为深棕色,细胞质淡染;染色阴性细胞胞核呈蓝色,细胞质未见染色。Sham组中,GRP78阳性细胞呈现少量表达,缺血再灌注后24 h,I/R组阳性细胞明显增多,两组比较差异具有统计学意义(P<0.01)。Caspase-12抗原阳性细胞呈棕黄色,以细胞质表达为主,在Sham组中微量表达,再灌注24 h后,I/R组阳性细胞较Sham组明显增多,两组比较差异具有统计学意义(P<0.01),见图3。

A、B:GRP78;C、D:Caspase-12;*:P<0.01,与Sham组比较
图4 Western blot检测GRP78和Caspase-12蛋白表达
2.6Western blot检测GRP78、Caspase-12蛋白表达 Sham组大鼠脑组织仅有少量GRP78、Caspase-12蛋白表达,I/R组24 h表达GRP78、Caspase-12蛋白较Sham组明显增加,两组比较差异有统计学意义(图4)。
3 讨 论
听皮层是大脑皮质的一个亚皮质区域,也是听觉系统的最高级中枢,是众多下调通路的源头,能在听觉系统的各个层面上影响神经信息处理。听觉皮质接受来自大脑中动脉的血供,皮层的功能也与缺血性损伤密切相关。听觉皮质神经元在缺血缺氧情况下容易发生细胞凋亡。课题组在前期的工作中采用夹闭双侧颈总动脉方法造成全脑缺血再灌注模型,观察了内质网应激反应及相关因子在听皮层的变化[7-8]。在本研究中,通过建立局灶性缺血再灌注模型,并通过神经功能评分、TTC染色证实模型制作成功。从形态学上可以观察到I/R损伤区细胞有明显的变化。对两组大鼠进行ABR测试发现,与Sham组相比,I/R组显示出ABR反应阈明显升高。因此笔者推断听皮质损伤可能是听力损伤早期阶段最重要的改变。
内质网是细胞内蛋白质合成以及修饰的主要场所,同时也是细胞内钙离子的储存库[9-11]。神经系统的内质网系统高度发达,对内外部环境的刺激如缺血缺氧、氧化应激及营养物质缺乏等非常敏感。当遇到各种有害刺激时,可以引起ERS,此时大量蛋白质在细胞内未正确折叠或错误折叠并大量堆积,从而导致钙离子紊乱。当这些蛋白质少量出现时,可以通过内质网相关降解途径进行清除。但大量出现并且超出了内质网的自身清除能力时,细胞内一系列级联反应通路被激活,促使具有保护作用的蛋白质生成,使细胞免受应激反应的损伤,维护机体的内环境稳定和生理平衡,称为非折叠蛋白反应(unfoldedproteinresponse,UPR)。但是如果严重而持续的ERS超过自身调节能力,则导致细胞凋亡和组织损伤[12-13]。
内质网应激相关性凋亡及凋亡相关蛋白近年来成为研究的热点。ERS诱导的UPR主要通过转录激活因子-6(activating transcription factor 6,ATF-6)、蛋白激酶样内质网激酶(phosphorylated extracellular signal regulated kinase,PERK)、肌醇依赖性蛋白激酶(inositol requiring enzyme 1,IRE1)3种感应蛋白介导的信号通路来实现[14]。GRP78被认为是ERS的经典标志物之一。在正常情况下与上述3个感应蛋白结合,封闭了它们的信号。当机体处于应激状态下,GRP78首先被释放出来与非折叠蛋白结合,达到阻止这些非折叠蛋白输出内质网的目的,进而对细胞起保护作用[15]。在本研究中,局灶性缺血再灌注24 h后GRP78表达明显上调提示,缺血再灌注启动了ERS过程。
Caspase-12途径是内质网应激引发凋亡的一个重要途径。正常情况下Caspase-12是ERS诱发凋亡的特异性介质,非ERS相关刺激诱导的细胞凋亡中不被激活。研究表明,抑制Caspase-12介导的ERS通路后,缺血再灌注后的神经细胞凋亡可以减少。对于Caspase-12基因缺陷大鼠,可以抵抗由于内质网应激引起的凋亡,对于其他死亡刺激并不能抵抗凋亡的发生[16-17]。在本研究中,局灶性脑缺血再灌注后,缺血侧听皮层凋亡阳性细胞明显增多,Caspase-12蛋白表达明显增高,提示内质网应激启动了相关凋亡途径,并进一步激活相关级联反应,导致神经元发生凋亡。
在脑缺血再灌注损伤过程中,内质网应激介导的凋亡途径可能是脑缺血损伤听皮层引发听力下降的可能机制之一。因此,如何抑制内质网应激及其相关凋亡途径,可为治疗伴有脑血管疾病的突发性耳聋提供了新的研究思路及研究方向。
[1]GAO H J,LIU P F,LI P W,et al.Ligustrazine monomer against cerebral ischemia/reperfusion injury[J].Neur Regen Res,2015,10(5):832-840.
[2]ISHIGE K,OSADA N,KOSUGE Y,et al.Involvement of endoplasmic reticulum stress in neurodegeneration after transient global ischemia-reperfusion[J].Nihon Yakurigaku Zasshi,2013,142(1):9-12.
[3]KIM H A,LEE B C,HONG J H,et al.Long-term prognosis for hearing recovery in stroke patients presenting vertigo and acute hearing loss[J].J Neurol Sci,2014,339(1/2):176-182.
[4]汪志伟,杨海弟,李永贺,等.盐酸椒苯酮胺对豚鼠耳蜗缺血再灌注损伤后细胞凋亡的影响[J].中国听力语言康复科学杂志,2015,13(2):117-119.
[5]李永贺,李威,陈浩,等.盐酸椒苯酮胺对豚鼠耳蜗缺血再灌注损伤的听力保护作用[J].听力学及言语疾病杂志,2013,21(6):603-607.
[6]施宁华,张志坚,许燕,等.全脑缺血再灌注后大鼠耳蜗组织内巢蛋白与神经生长因子的表达[J].江苏大学学报,2011,21(1):5-8.
[7]吕哲,张颖,王团,等.豚鼠脑缺血再灌注后听皮层内质网应激相关因子的表达及意义[J].听力学及言语疾病杂志,2016,24(4):377-381.
[8]路虹,王团,徐鸥,等.内质网应激反应对豚鼠脑缺血再灌注后听皮质区损伤的作用[J].中国耳鼻咽喉头颈外科,2014,21(5):249-254.
[9]WU CX,LIU R,GAO M,et al.Pinocembrin protects brain against ischemia/reperfusion injury by attenuating endoplasmic reticulum stress induced apoptosis[J].Neurosci Lett,2013,546(1):57-62.
[10]QIU B,HU S D,LIU L B,et al.CART attenuates endoplasmic reticulum stress response induced by cerebral ischemia and reperfusion through upregulate BDNF synthesis and secretion[J].Biochem Biophys Res Commun,2013,436(4):655-659.
[11]JOHNSON G G,WHITE M C,WU J H,et al.The deadly connection between endoplasmic reticulum,Ca2+,protein synthesis,and the endoplasmic reticulum stress response in maliongnant glioma cells[J].Neuro Oncol,2014,16(8):1086-1099.
[12]NISHITOH H.CHOP is a multifunctional transcription factor in the ER stress response[J].J Biochem,2012,151(3):217-219.
[13]SU Y,LI F.Endoplasmic reticulum stress in brain ischemia[J].Int J Neurosci,2016,126(8):681-691.
[14]KREBS J,AGELLON L B,MICHALAK M.Ca2+homeostasis and endoplasmic reticulum (ER) stress:An integrated view of calcium signaling[J].Biochem Biophys Res Commun,2015,460(1):114-121.
[15]ZHU G,LEE A S.Role of the unfolded protein response,GRP78 and GRP94 in organ homeostasis[J].J Cell Physiol,2015,230(7):1413-1420.
[16]XU X,LIU T,ZHANG A,et al.Reactive oxygen species-triggered trophoblast apoptosis is initiated by endoplasmic reticulum stress via activation of caspase-12,CHOP,and the JNK pathway in toxoplasma gondii infection in mice[J].Infect Immun,2012,80(6):2121-2132.
[17]林岩,肖薇,金莉,等.内质网应激介导白藜芦醇对心肌细胞肥大的保护作用[J].中国药理学通报,2014,30(12):1721-1725.
猜你喜欢
解放军医学杂志(2021年12期)2022-01-18 03:53:24
现代临床医学(2021年1期)2021-01-26 00:55:52
中国现代医药杂志(2020年3期)2020-05-08 04:33:08
中国生物医学工程学报(2019年6期)2019-07-16 07:52:48
安徽医科大学学报(2016年12期)2017-01-15 14:21:55
中外医疗(2016年15期)2016-12-01 04:25:39
中国组织化学与细胞化学杂志(2016年3期)2016-02-27 11:15:28
中国当代医药(2015年33期)2015-03-01 02:09:08
西安交通大学学报(医学版)(2014年5期)2014-06-23 16:28:26
西安交通大学学报(医学版)(2014年5期)2014-06-23 16:28:26
