
云南三个地方猪种肌肉脂肪酸组成分析*
2018-01-24 11:02杨洁鸿李星润严达伟董新星
家畜生态学报 2018年1期
杨洁鸿,李星润,胡 伟,严达伟,董新星*,苟 潇*
(1.云南农业大学 动物技术学院,云南 昆明 650201;2.大理农林职业技术学院 动物科学系,云南 大理 671003)
猪肉作为我国传统肉食品,被广泛应用在罐头食品、冷冻食品以及零食等加工中。猪肉中瘦肉的脂肪含量高达25%~30%,远高于其他肉质产品。脂肪是生命体重要的营养物质,由甘油和脂肪酸构成。在机体内,脂肪一部分被氧化分解为机体供能,另一部分以脂肪酸的形式被机体吸收利用。脂肪酸根据人类需求不同可以分为必需脂肪酸和非必需脂肪酸[1],猪肉中含有大量的必需脂肪酸,可以为人类提供必要的营养物质。脂肪酸根据分子结构的不同可以分为饱和脂肪酸、单不饱和脂肪酸、多不饱和脂肪酸,其中单不饱和脂肪酸可以通过调节低密度脂蛋白氧化敏感性来降低心血管疾病的发生率;多不饱和脂肪酸具有抗炎、提高免疫力、促进血液循环、减少动脉粥样硬化的发生几率等功能[2]。然而大多猪种脂肪酸中大部分为饱和脂肪酸,饱和脂肪酸的过量摄入会导致机体高血脂、高胆固醇症的发生[3]。饱和脂肪酸具有稳定的分子结构,不会随高温等外界条件改变,这些性状阻碍了猪肉作为理想的营养产品的潜力。
云南地处西南边陲,地貌复杂,自然生态环境多样,具有丰富的遗传资源。本研究选取滇南小耳猪,撒坝猪,大河乌猪作为研究肉质脂肪酸组成的主要材料。滇南小耳猪属于华南型猪种,早熟易肥,肉质优良,口感肥而不腻,鲜香嫩糯;撒坝猪是乌金猪的一个重要类型,耐粗饲,易催肥肉质好,大理石纹发达,肌纤维较细,适宜制作火腿和腌肉;大河猪是腌制“云腿”的传统原料猪,与杜洛克杂交后育成大河乌猪,大河乌猪具有较高生长速度、产肉性能和饲料转化率,且肉香味美[4]。本研究旨在通过对云南本地猪(滇南小耳猪,撒坝猪,大河乌猪)背最长肌脂肪酸成分的分析,比较各类脂肪酸差异,为探究云南本地猪肉营养价值,发掘优秀猪种遗传资源及云南本土猪种的推广养殖提供一定的理论基础。
1 材料与方法
1.1 试验材料
本研究所采用试验猪共49头,其中撒坝猪8头(4♂,4♀),来自云南省昆明市东川区撒坝某猪资源保种场,屠宰猪只日龄均为200日龄左右,体重为85~90 kg;大河乌猪24头(12♂,12♀),来自云南省曲靖市某猪场,屠宰猪只日龄均为180日龄左右,体重为80~90 kg;滇南小耳猪17头(8♂,9♀),云南省西双版纳滇南小耳猪某保种场,屠宰猪只日龄均为300日龄左右,体重为40~45 kg。各猪种饲养方式均采用全价配合饲料加适当青绿饲料混合饲喂,圈养。
1.2 指标测定
将试验猪进行屠宰,取左半胴体倒数第3~4腰椎处的背最长肌100 g,迅速冰冻,置于冻存盒中立即送广州市农业标准与检测中心。制备各脂肪酸标准贮备液并绘制脂肪酸浓度标准曲线。取5 g肉样解冻匀浆干燥后取0.5 g干样,加苯与石油醚(1: 1)混合液,浸提后加氢氧化钾-甲醇溶液(0.4 mol/l)进行甲酯化,取上清液过滤后供气相色谱分析。与标准样作同样的测定,由标准曲线查得各脂肪酸的浓度,用面积归一法测定脂肪酸组成[5]。每个样品平行测定三次,取平均值为最后测定结果。测定撒坝猪8头,大河乌猪24头,滇南小耳猪17头,共49头。
1.3 数据处理
所有数据采用SAS 9.0进行统计分析,各品种背
最长肌数据作单因素方差分析,所用数学模型为:
yij=μ+ai+ eij
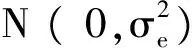

2 结果与分析
2.1 脂肪酸的组成
本研究测定大河乌猪、撒坝猪、滇南小耳猪背最长肌中的脂肪酸含量,根据样品进行气相色谱分析得到的峰面积与标准样品的峰面积相比见表1。从表1可知,3种猪肌肉样品均由10种饱和脂肪酸(SFA)以及9种不饱和脂肪酸(UFA)组成,包括4种单不饱和脂肪酸(MUFA)和5种多不饱和脂肪酸(PUFA)。饱和脂肪酸包括辛酸(C8:0)、癸酸(C10:0)、十二酸(C12:0)、十四酸(C14:0)、十五酸(C15:0)、十六酸(C16:0)、十七酸(C17:0)、十八酸(C18:0)、十九酸(C19:0)、二十酸(C20:0),单不饱和脂肪酸包括十六碳一烯酸(C16:1n-14)、十七碳一烯酸(C17:1n-7)、油酸(C18:1n-9)、二十碳一烯酸(C20:1n-9),多不饱和脂肪酸包括亚油酸(C18:2n-6)、亚麻酸(C18:3n-3)、二十碳二烯酸(C20:2n-6)、二十碳三烯酸(C20:3n-3)、二十碳四烯酸(C20:4n-6)。

表1 3种猪背最长肌的脂肪酸含量Table 1 Fatty acid contents in longissimus dorsi of 3 breeds
注:“*”脂肪酸数据为不同猪种的背最长肌的算术平均数,同行中小写字母不同表示差异显著(P<0.05) ,大写字母不同表示差异极显著(P<0.01)。
Note: “*” fatty acids data is the arithmetic average in the longissimus of the 3 breeds. In the same line,different lowercase letter superscripts indicate significant difference (P<0.05) and different capital letter superscripts indicate extremely significant difference (P<0.01) .
2.2 饱和脂肪酸的含量比较
从表1可知,撒坝猪背最长肌的饱和脂肪酸含量极显著低于大河乌猪和滇南小耳猪的含量(P<0.01),大小依次为大河乌猪>滇南小耳猪>撒坝猪。在饱和脂肪酸中,十六酸(C16:0)的含量最高,其中大河乌猪和撒坝猪的含量极显著高于滇南小耳猪(P<0.01);其次是十八酸(C18:0)和十四酸(C14:0),在十八酸(C18:0)中,3种猪之间均差异极显著(P<0.01),从大到小为滇南小耳猪>大河乌猪>撒坝猪;在十四酸(C14:0)中,滇南小耳猪差异极显著高于大河乌猪(P<0.01),且撒坝猪与大河乌猪、滇南小耳猪差异均不显著(P>0.05);剩余7种饱和脂肪酸含量很少。由此可见,所测定的10种饱和脂肪酸中,主要差异存在于C14、C16和C18系列中,其中最突出的是C16:0。
2.3 不饱和脂肪酸含量的比较
从表1可知,本研究中所有猪种饱和脂肪酸的含量均低于不饱和酸的含量,撒坝猪不饱和脂肪酸的含量均显著高于大河乌猪和滇南小耳猪(P<0.01),大小依次为撒坝猪>滇南小耳猪>大河乌猪。在单不饱和脂肪酸中,滇南小耳猪含量显著低于大河乌猪和撒坝猪(P<0.01),大小依次为撒坝猪>大河乌猪>滇南小耳猪;其中,油酸(C18:1n-9)的含量最高,同样是滇南小耳猪含量显著低于大河乌猪和撒坝猪(P<0.01)。在多不饱和脂肪酸中,滇南小耳猪含量极显著高于撒坝猪和大河乌猪(P<0.01),大小依次为滇南小耳猪>撒坝猪>大河乌猪,其中含量最高的是亚油酸(C18:2n-6),其次为亚麻酸(C18:3n-3)、二十碳二烯酸(C20:2n-6)、二十碳四烯酸(C20:4n-6)和二十碳三烯酸(C20:3n-3)。亚油酸(C18:2n-6)、亚麻酸(C18:3n-3)、二十碳四烯酸(C20:4n-6)是人体必需脂肪酸,其中滇南小耳猪的这3种含量均极显著高于大河乌猪和撒坝猪(P<0.01)。
3 讨 论
3.1 饱和脂肪酸营养价值分析
在饱和脂肪酸中,十六酸(C16:0)和十八酸(C18:0)远大于其他饱和脂肪酸,含量分别为22.04%~26.29%和12.75%~17.80%。其他的地方猪种的十六酸(C16:0)和十八酸(C18:0)的含量也占脂肪酸的很大比例,如莆田黑猪、杜莆、大莆,含量为24.08%~27.57%和12.75%~13.45%[7]。十六酸(C16:0)能够促进婴幼儿的营养吸收,且是人体重要的产能脂肪酸,十八酸(C18:0)可减轻胆汁淤积症引起的肝损伤[8],所以对人体都很重要[8]。有研究表明,猪肉嫩度、多汁性及风味口感与肌肉中饱和脂肪酸、单不饱和脂肪酸的高含量有直接关系[10-11]。大河乌猪、撒坝猪和滇南小耳猪饱和脂肪酸和单不饱和脂肪酸总的含量分别为92.97%、92.16%和84.81%,其中大河乌猪、撒坝猪饱和脂肪酸和单不饱和脂肪酸总的含量均大于杜洛克(88.77%)和大约克(87.76%)的含量[7],说明云南地方猪种大河乌猪、撒坝猪的肉质风味更好。
3.2 不饱和脂肪酸营养价值分析
研究表明,单不饱和脂肪酸能够降低心脏病的发生几率,对心脏有保护作用,同时还有降低血糖、调节血脂、降低胆固醇、防止记忆减退等作用[12]。大河乌猪和撒坝猪单不饱和脂肪酸的含量分别为49.37%和50.58%,比杜洛克(49.02%)和大约克(48.87%)的含量高。单不饱和脂肪酸中,油酸(C18:1n-9)的含量最高,其可以调整高中低3种密度胆固醇的浓度,并且能够提供大量能量[13]。大河乌猪和撒坝猪油酸(C18:1n-9)的含量分别为49.37%和50.58%,比莆田黑猪(42.31%)、杜莆(42.5%)、大莆(43.06%)、杜洛克(42.04%)和大约克(42.46%)的含量都要高[7]。撒坝猪单不饱和脂肪酸含量远高于杜浙猪、杜长大[14]。研究证实,多不饱和脂肪酸具有对多巴胺神经元和视网膜的保护作用、有助于大脑活力的提升、促进胎儿和婴幼儿大脑发育[15]。其中亚油酸(C18:2n-6)、亚麻酸(C18:3n-3)二十碳四烯酸(C20:4n-6)是人体必需脂肪酸。滇南小耳猪多不饱和脂肪酸、亚油酸(C18:2n-6)、亚麻酸(C18:3n-3)和二十碳四烯酸(C20:4n-6)的含量均高于莆田黑猪、杜莆、大莆、杜洛克和大约克的含量;撒坝猪多不饱和脂肪酸含量以及亚油酸(C18:2n-6)和二十碳四烯酸(C20:4n-6)的含量均大于莆田黑猪、杜莆、大莆含量;大河乌猪多不饱和脂肪酸和亚油酸(C18:2n-6)的含量均高于杜莆的含量,且二十碳四烯酸(C20:4n-6)的含量高于杜洛克的含量[7]。滇南小耳猪多不饱和脂肪酸含量高于新吉林黑猪、巴克夏猪、大白猪以及杜浙猪、杜长大[14,16]。
不同品种猪的肌内脂肪含量有所不同,脂肪酸的组成也不尽相同[17-19],究其原因是各品种的遗传背景差异。近年来,影响肌内脂肪的候选基因,如心脏脂肪酸结合蛋白(H-FABP)基因、脂肪细胞脂肪酸结合蛋白(A-FABP)基因等已在不同品种猪群中进行了充分研究。激素敏感脂肪酶(HSL)基因、脂蛋白脂肪酶(LPL)基因、肥胖(OB)基因和过氧化物酶增殖激活受体(PPAR)等被认为是影响猪脂肪沉积的重要候选基因。影响脂肪酸成分的QTL也正在被逐步定位,目前已发现85个,占据了至少5%的基因组。伊比利亚猪和长白猪杂交群体中分别在4号、8号、10号和12号染色体发现显著影响亚油酸(C18:2n-6)、十六酸(C16:0)、十六碳一烯酸C16:1(n-9)、十四酸(C14:0)和亚麻酸(C18:3n-3)含量的QTL[20-21]。长白公猪和约克夏母猪的杂交群体中,在1号和18号染色体分别检测到影响亚油酸(C18:2n-6)和十四酸(C14:0)含量的QTL[22]。日本野公猪和大白母猪的杂种后代中,在9条染色体上定位了影响7种脂肪酸成分的QTL[23]。杜洛克公猪和大白母猪杂交群体中,在5条染色体上定位了9个显著影响脂肪酸的QTL[24]。各品种脂肪酸组成的不同跟遗传背景有很大关系,随着对影响脂肪酸的QTL的深入研究,可深入揭示各品种脂肪酸组成差异的遗传原因。
[1] 陈银基,鞠兴荣,周光宏.饱和脂肪酸分类与生理功能[J].中国油脂,2008,33(3):35-39.
[2] UDHAYABANU T, MANOLE A, RAJESHWARI M, et al. Riboflavin responsive mitochondrial dysfunction in neurodegenerative diseases[J]. Journal of Clinical Medicine, 2017, 6(5): E65.
[3] LI Yuwen, PARK J, WU Yin, et al. Identification of AMPK activator from twelve pure compounds isolated from Aralia Taibaiensis: implication in antihyperglycemic and hypolipidemic activities[J]. The Korean Journal of Physiology & Pharmacology : Official Journal of the Korean Physiological Society and the Korean Society of Pharmacology, 2017, 21(3): 279-286.
[4] 连林生.云南地方猪种的选育及其改良途径的研究[J].猪业科学,2008,25(4):102-104.
[5] 国家卫生和计划生育委员会.国家食品药品监督管理总局.食品安全国家标准食品中脂肪酸的测定:BG5009.168-2016[S].北京:中国标准出版社,2017.
[6] 鲁绍雄,连林生.SAS 统计分析系统在畜牧科学中的应用[M].昆明:云南科技出版社,2003:66.
[7] 吴妹英,曹长贤,张力,等.不同品种猪肌肉脂肪酸和氨基酸含量[J].福建农林大学学报(自然科学版),2009,38(2):166-170.
[8] PAN P H, LIN S Y, OU Y C, et al. Stearic acid attenuates cholestasis-induced liver injury[J]. Biochemical and Biophysical Research Communications, 2010, 391(3): 1 537-1 542.
[9] 商允鹏,生庆海,王贞瑜,等.三酰甘油Sn-2位上棕榈酸生理功能及研究概况[J].中国粮油学报,2010,25(10):119-123.
[10] CAMERON N D, ENSER M B. Fatty acid composition of lipid in Longissimus dorsi muscle of Duroc and British Landrace pigs and its relationship with eating quality[J]. Meat Science, 1991, 29(4): 295-307.
[11] 徐秋良,吴运香,张长兴,等.畜禽肉嫩度及其影响因素[J].家畜生态学报,2010,31(6):100-103.
[12] 张伟敏,钟耕,王炜.单不饱和脂肪酸营养及其生理功能研究概况[J].粮食与油脂,2005(3):13-15.
[13] 吴时敏.功能性油脂[M].北京:中国轻工业出版社,2001.
[14] 门小明,邓波,徐子伟,等.两种杂交商品猪冷却肉常规养分含量、蛋白质溶解性及脂肪酸组成的比较[J].食品工业科技,2012,33(17):361-364.
[15] 崔和平,郭兴凤.多不饱和脂肪酸对人体神经系统保健作用研究进展[J].河南工业大学学报(自然科学版),2012,33(3):101-106.
[16] 李娜,张树敏,李兆华,等. 三个猪种肌肉营养成分分析[J]. 黑龙江畜牧兽医,2015(20):61-63.
[17] 陶立,金邦荃,尹晴红,等.F1代猪体脂,肌肉脂肪和游离脂肪酸组分的分析[J].江苏农业学报,2001,17(2):101-103.
[18] 吴德,杨凤,周安国,等.不同比例梅山猪血缘生长育肥猪肉质及肌纤维组织学特性研究[J].四川农业大学学报,2001,19(3):252-255.
[19] 刘宗华,张牧,邢军.杂交对小梅山猪肌肉营养成分的影响[J].养猪,2002(4):31-32.
[21] CLOP A, OVILO C, PEREZ-ENCISO M, et al. Detection of QTL affecting fatty acid composition in the pig[J]. Mammalian Genome : Official Journal of the International Mammalian Genome Society, 2003, 14(9): 650-656.
[22] LEE C, CHUNG Y, KIM J H. Quantitative trait loci mapping for fatty acid contents in the backfat on porcine chromosomes 1, 13, and 18[J]. Molecules and Cells, 2003, 15(1): 62-67.
[23] NII M, HAYASHI T, TANI F, et al. Quantitative trait loci mapping for fatty acid composition traits in perirenal and back fat using a Japanese wild boar x Large White intercross[J]. Animal Genetics, 2006, 37(4): 342-347.
[24] SANCHEZ M P, IANNUCCELLI N, BASSO B, et al. Identification of QTL with effects on intramuscular fat content and fatty acid composition in a Duroc x Large White cross[J]. BMC Genetics, 2007, 8(1): 55.
猜你喜欢
猪业科学(2022年10期)2022-11-03
社会科学战线(2022年2期)2022-03-16
猪业科学(2021年6期)2021-08-12
云南医药(2021年3期)2021-07-21
Asian Herpetological Research(2021年2期)2021-06-30
猪业科学(2021年3期)2021-05-21
水产科学(2018年2期)2018-03-27
水产科学(2017年3期)2017-12-18
中国粮油学报(2016年5期)2016-01-23
中国粮油学报(2016年5期)2016-01-23
