
山东栒子光合特性的研究
2017-05-17 11:59杨海平姚树建赵永军刘建华
山东林业科技 2017年4期
杨海平 ,张 锋 ,姚树建 ,赵永军 ,刘建华 ,韩 义 ,王 磊 *
(1.山东省林木种质资源中心,济南 250014;2.招远市林业局,招远 265400)
由于近几十年来我国人口的快速增长及经济的高速发展,对野生植物生境的破坏、掠夺式的开发利用和环境污染等原因,致使许多野生植物的生存面临严重威胁。山东栒子 (Cotoneaster schantungensisKlotz.)属于蔷薇科栒子属植物,为山东特有种,仅分布于济南东南部山区,因其种群数量少,已处濒危状态,相关研究资料少,主要集中在种群分布与组成以及扦插繁育方面[1,2],对其光合特性方面研究尚未见报道。为更好保护和利用这一宝贵资源,我们以其常见的近缘种西北栒子(Cotoneaster zabeliiSchneid.)为参照,比较分析它们光合特性的差异,讨论山东栒子对自然环境的适应能力,为其进一步开发利用、扩大栽培提供科学依据和理论指导。
1 材料与方法
1.1 试验地概况
本研究试验地点设在山东林木种质资源中心枣园种质资源库(以下简称枣园库)。枣园库地处山东省章丘市枣园镇,北纬 36°4′,东经 117°33′。土层厚度大多超过120cm,土壤类型为褐土,呈中性至微碱性。该地属暖温带季风区大陆性气候,四季分明,全年平均气温12.9℃,年平均大于10℃的积温4584℃,全年无霜期192天。平均年日照时数2638.5h,平均降雨量628mm,降雨集中在6-8月份[3]。
1.2 试验材料
供试材料采集于济南佛峪地区的山东栒子及西北栒子的种子,通过播种育苗繁殖获得苗木。
1.3 试验方法
试验在2013年7-9月份选择晴朗无风天气进行。采用LI-6400XT便携式光合测定系统(Li-COR,美国),选择无病虫害,长势良好一致的山东栒子和西北栒子各3株,选取采光良好的成熟叶片进行测定,每个叶片记录3次。测定植株净光合速率Pn,同时记录光合有效辐射PAR、蒸腾速率Tr、胞间CO2浓度Ci和气孔导度Gs等指标。
1.3.1 光合日变化的测定
测定使用透明叶室采用自然光源,以大气中CO2为碳源,从6:00-18:00每隔2h对山东栒子和西北栒子进行一次光合测定。
1.3.2 光响应曲线(Pn-PAR)的测定
测定使用Li-6400-02B红蓝光源,PAR在0~2000μmoL·m-2·s-1范围内设定16个梯度(2000,1800,1600,1400,1200,1000,800,600,400,200,150,100,80,50,20,0μmoL·m-2·s-1), 测量前进行一定时间的光诱导,CO2浓度控制在 400 μmoL·moL-1。测定时间为上午9:00-11:00。
1.3.3 二氧化碳响应曲线(Pn-CO2)的测定
测定使用Li-6400-02B红蓝光源,PAR设定为800μmoL·m-2·s-1,CO2浓度值的设置顺序为 400,300,200,150,100,70,50,0,400,400,600,800,1000,1200,1500μmoL·moL-1,共 15 个梯度。测定时间为上午 9:00-11:00。
1.4 数据处理
采用SPSS19.0、Excel2003和光合计算软件3.3对数据进行方差分析和响应曲线的拟合。
1.4.1 光响应曲线拟合模型
采用光饱和点与实测值较为接近的直角双曲线的修正模型作为计算依据[4,5]。

进行Pn-PAR曲线拟合时,式中Pn为净光合速率,Pnmax为最大净光合速率,LSP为光饱和点,LCP为光补偿点,α为光响应曲线的初始斜率(无量纲),即表观光合量子效率AQY;I为光合有效辐射(PAR),β和γ为系数;Rd为暗呼吸速率。
1.4.2 CO2响应曲线拟合模型
采用饱和胞间CO2浓度与实测值较为接近的直角双曲线的修正模型作为计算依据[6]。

进行Pn-CO2曲线拟合时,式中Pn为净光合速率,P*nmax为最大净光合速率,Cisat表示植物的饱和胞间CO2浓度,CCP表示CO2补偿点,Ci为胞间CO2浓度,Rp为光呼吸速率,a为植物光合作用对CO2响应曲线的初始斜率,也称为初始羧化效率(无量纲),b和c为系数。
2 结果与分析
2.1 光合指标日变化
图 1(a、b、c、d)分别给出山东栒子和西北栒子净光合速率 Pn、气孔导度 Gs、胞间CO2浓度 Ci和蒸腾速率Tr日变化情况:由图a可以看出,山东栒子和西北栒子净光合速率日变化曲线均接近单峰型,上午10:00净光合速率均达最大值,之后开始下降并在一段时间内(12:00-14:00)维持平稳,在14:00-16:00随光辐射降低净光合速率迅速降低,16:00-18:00净光合速率继续下降并出现负值说明光合作用积累的有机物已不能满足植物的呼吸消耗。山东栒子净光合速率日变化平均值要高于西北栒子,说明山东栒子对光的适应性更好、利用率更高,单位时间积累有机物的量更大。图b和图d中两种植物气孔导度和蒸腾速率日变化呈现相同趋势:西北栒子气孔导度和蒸腾速率在一天中的变化相对平稳,山东栒子的气孔导度和蒸腾速率在一天中明显高于西北栒子且呈现升高-维持-降低趋势。图c中,山东栒子和西北栒子胞间CO2均呈现先降后升趋势,并在10:00时达到最低值,分析原因可能是此时植物光合作用强造成CO2大量消耗。综合分析,在外界环境基本一致条件下,山东栒子气孔导度明显高于西北栒子,使山东栒子与外界环境气体交换更容易和频繁,进而获得更多光合作用所需的CO2,同时,气体交换量大又使山东栒子植株蒸腾作用更强,推测其在生长过程中比西北栒子需水量更大。

图1 Pn及各气体交换参数的日变化Fig.1 The diurnal change of Pn and gas exchange parameters a:净光合速率日变化;b:气孔导度日变;c:胞间CO2浓度日变;d:蒸腾速率日变化Tr
2.2 光响应曲线(Pn-PAR)
利用直角双曲线修正模型对山东栒子和西北栒子光响应曲线进行拟合,由图2可以看出山东栒子和西北栒子Pn-PAR曲线拟合非常好,拟合系数分别为0.9992和0.996。图2中山东栒子和西北栒子Pn在一定范围内随PAR增大而逐渐升高,当光PAR继续增大并达到一定值时植株Pn趋于平稳甚至出现下降趋势。 由图 2可知,PAR<200μmoL·m-2s-1时,山东栒子Pn值小于西北栒子,PAR>200μmoL·m-2s-1时,山东栒子Pn值明显大于西北栒子,表明山东栒子利用弱光的能力弱于西北栒子,但利用强光的能力明显较强。结合直角双曲线修正模型计算山东栒子和西北栒子 Pnmax,Rd,LSP,LCP,AQY 等相关光合生理参数,结果如表1。山东栒子Pnmax,LSP,LCP值均高于西北栒子,说明山东栒子比西北栒子利用光的能力强,有利于有机物的积累。AYQ反应植物利用弱光的能力,山东栒子 AYQ为0.0372 μmoL CO2·μmoL-1photons低于西北栒子 AYQ 值 0.0522μmoL CO2·μmoL-1photons,说明山东栒子较西北栒子利用弱光的能力弱。山东栒子Rd值高于西北栒子,说明其暗呼吸速率较大,不利于有机物的积累。

图2 光响应曲线Fig.2 The light response curves
2.3 二氧化碳响应曲线(Pn-CO2)
利用直角双曲线修正模型对山东栒子和西北栒子CO2响应曲线进行拟合,由图3可以看出山东栒子和西北栒子Pn-CO2曲线拟合非常好,拟合系数为0.9973和0.9981。Pn-CO2曲线与Pn-PAR曲线变化趋势相同,即在一定CO2浓度范围内山东栒子和西北栒子净光合速率随CO2浓度增加而增大,随着CO2浓度继续增大,净光合速率增加趋于平稳甚至呈降低趋势。在CO2浓度较低时 (约小于100μmoL·moL-1),山东栒子比西北栒子净光合速率低,随着CO2浓度增大,相同CO2浓度条件下山东栒子净光合速率始终高于西北栒子,说明山东栒子对CO2利用率较高,利于有机物的积累。结合直角双曲线修正模型,计算出Pn-CO2响应曲线的P*nmax,Cisat,CCP等参数,山东栒子P*nmax,CCP均高于西北栒子,说明山东栒子CO2利用率较高,山东栒子Cisat虽低于西北栒子,但环境中CO2浓度一般在400μmoL·moL-1左右,因此不具备实际意义。
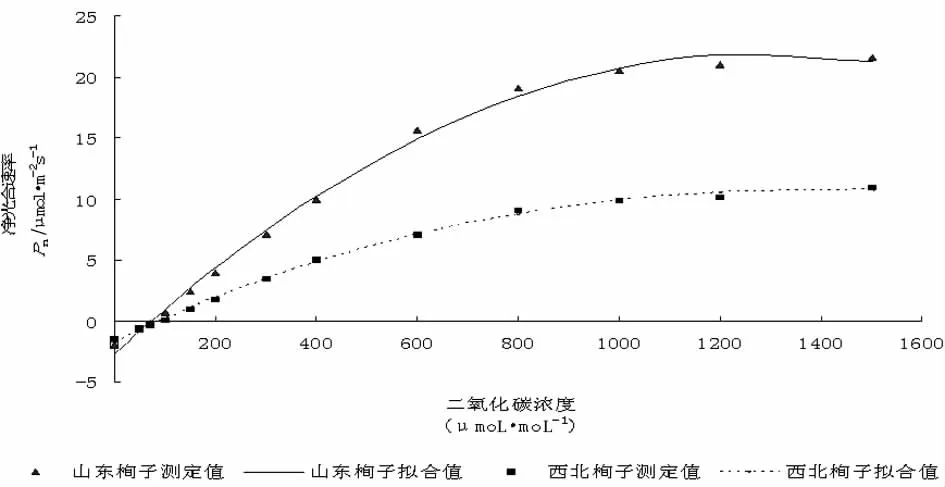
图3 CO2响应曲线Fig.3 The CO2response curves

表1 光响应曲线参数分析表Tab.1 The parameter analyses of the light response curves

表2 CO2响应曲线参数分析表Tab.2 The parameter analyses of the CO2response curves
3 结论与讨论
植物光合作用是一个连续的过程,通过测定植物白天的各项光和参数变化情况能够反映植物对环境的适应及对环境中光合条件的利用情况。试验结果显示,山东栒子和西北栒子净光合速率日变化均接近于单峰型,净光合速率的峰值出都现在10:00,在外界环境相近条件下试验比较山东栒子和西北栒子净光合速率、气孔导度、胞间CO2浓度和蒸腾速率日变化情况。可以发现,山东栒子一天中气孔导度明显高于西北栒子,说明植株与环境进行更多的气体交换,因此山东栒子一天中蒸腾作用和胞间CO2浓度均明显高于西北栒子。这些影响因子在植物光合作用上的综合表现为山东栒子净光合速率的日平均值高于西北栒子,说明山东栒子对外界环境适应性更好。
[1]屈素青,刘丹,解孝满,等.山东栒子群落组成与结构特征分析[J].安徽农业科学.2012,40(3):1426-1427,1430.
[2]李朝晖,李文清,解孝满,等.珍稀濒危植物山东栒子扦插技术研究[J].山东林业科技.2014,(2):69-71.
[3]刘启虎,鲁仪增,李文清,等.假植对北美鹅掌楸生长量的影响分析研究[J].山东林业科技.2009,(3):56-58.
[4]Ye Z.P,Yu Q.Comparison of new and several classical models of photosynthesis in response to irradiance[J].Journal of Plant Ecology.2008,(32):1356–1361.
[5]Ye Z.P,Yu Q.A coupled model of stomatal conductance and photosynthesis for winter wheat[J].Photosynthetica,2008.(46):637–640.
[6]Ye Z.P,Yu Q.Comparison of photo-synthetic response to intercellular CO2and air CO2[J].Journal of Ecology.2009,(28),2233-2238.
猜你喜欢
东北林业大学学报(2022年4期)2022-05-16
农业工程学报(2022年3期)2022-04-16
湖北农业科学(2021年13期)2021-08-04
新疆农业科技(2021年2期)2021-07-03
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
中学生物学(2020年12期)2020-03-30
中国农业科技导报(2020年12期)2020-03-15
