
低氧联合NaCl胁迫下外源Ca2+对发芽苦荞γ-氨基丁酸富集的影响
2017-04-13 02:34朱云辉郭元新杜传来丁志刚
中国粮油学报 2017年1期
朱云辉 郭元新 杜传来 丁志刚 叶 华
(安徽科技学院食品药品学院,凤阳 233100)
低氧联合NaCl胁迫下外源Ca2+对发芽苦荞γ-氨基丁酸富集的影响
朱云辉 郭元新 杜传来 丁志刚 叶 华
(安徽科技学院食品药品学院,凤阳 233100)
采用在通气的培养液中添加外源Ca2+培养的方式,研究了低氧联合NaCl胁迫下外源Ca2+对发芽苦荞GABA富集的影响。低氧联合NaCl胁迫对苦荞的芽长有抑制作用,GAD活力和GABA的含量随着NaCl浓度的增大均呈先上升后下降的趋势,在10 mmol/L NaCl溶液处理时均达到最高值,分别是单纯低氧胁迫的1.65倍和1.10倍;在低氧联合NaCl胁迫的基础上添加低浓度的Ca2+可缓解NaCl胁迫对苦荞芽长的抑制作用,添加3 mmol/L Ca2+处理可使GAD活力和GABA的富集量达到最大,分别为91.44 U/g FW和492.31 μg/g DW,分别是低氧联合NaCl胁迫的1.12倍和1.16倍;低氧联合NaCl胁迫下添加Ca2+螯合剂和通道阻断剂后发芽苦荞芽长显著降低,GAD活力显著下降,GABA富集量减少。外源Ca2+可降低低氧联合NaCl胁迫对苦荞芽长的抑制作用,并可通过提升GAD活力,增强发芽苦荞GABA的富集。
低氧胁迫 NaCl胁迫 Ca2+处理 发芽苦荞 GABA富集
芽类食品是东方人重要的蔬菜,已有2 000多年的历史。近年来,豆芽、苜蓿芽等芽类食品作为一种异域的健康和时尚产品,风靡欧美市场[1-2]。苦荞(Fagopyrum Tataricum)芽菜是中国特有的药食两用蔬菜,已有悠久的食用历史却一直没有得到应有的重视[3-5]。苦荞麦属蓼科作物,其丰富的蛋白质、脂肪、维生素、矿物质及多酚类物质含量都普遍高于大米、小麦、玉米等大宗粮食作物[6],特别是苦荞中含有禾谷类粮食所没有的卢丁等黄酮类物质[7]苦荞独特的降血压、降血脂和降血糖功能逐渐受到广大消费者们的青睐。近年来的研究表明,通过控制种子的发芽条件,可以富集γ-氨基丁酸(γ-aminobutyric acid,GABA)等植物中原来含量低或不具有的功能性成分[8-10]。
GABA是一种四碳非蛋白质氨基酸,广泛存在于真核和原核生物中,具有降血压、改善心脑机能及缓解疼痛和焦虑等作用,开发富含GABA的食品备受关注[11-12]。植物中GABA合成主要来自GABA支路中由谷氨酸脱羧酶(GAD,EC 4.1.1.15)催化的不可逆的α-谷氨酸脱羧反应[13]。植物受到冷激、干旱、盐胁迫和低氧等胁迫时会强烈刺激其GAD活性,使GABA大量积累[8,14-15],其中低氧胁迫富集GABA最为快速有效[16-17],近年来的研究还将低氧胁迫同NaCl胁迫结合起来,对GABA的富集起到了叠加效应[9,18]。GAD是一种Ca2+/钙调蛋白(CaM)依赖型的酶,具有一个钙调蛋白结合区[19],因此在培养过程中添加外源Ca2+可提高GAD活力,继而富集GABA。韩永斌等[20]将糙米放入通气的培养液中并向其加入一定浓度的Ca2+,发现GABA含量显著增加。目前,研究者们对苦荞发芽的研究多集中在黄酮类物质上[21-22],有关苦荞富集GABA的报道较少,利用低氧联合NaCl胁迫下添加Ca2+处理富集GABA的研究还鲜见报道。
本试验在低氧胁迫的基础上,通过添加NaCl、Ca2+和Ca2+抑制剂处理,研究其对发芽苦荞的生长、GAD活力和GABA富集的影响,并对相关作用机制做初步探讨,旨在为富含GABA的苦荞功能性芽菜食品的开发提供参考。
1 材料与方法
1.1 材料与试剂
苦荞(榆6-21):2014年秋产自中国内蒙古自治区乌兰察布市,于-20℃贮藏于冰箱;GABA标品(>99.9%)、对二甲氨基苯磺酰氯(DABS-CI,99%):美国Sigma公司。
1.2 仪器与设备
LHP-160型智能恒温恒湿培养箱:上海三发科学仪器有限公司;Agilent 1200液相色谱仪:安捷伦公司;KDC-160HR高速冷冻离心机:合肥科大创新股份有限公司;真空冷冻干燥系统:美国Labconce公司;LZB-3型空气流量计:江苏余姚工业自动化仪表厂。
1.3 试验方法
1.3.1 苦荞发芽试验
正常发芽:取30 g苦荞种子用去离子水浸泡清洗后,用1%的次氯酸钠溶液消毒15 min,然后用去离子水冲洗至pH值中性,在去离子水中30℃浸泡4 h后,放入铺有2层滤纸的φ9 cm培养皿中,在生化培养箱中30℃避光培养,培养期间每8 h左右喷去离子水1次,以保持发芽种子湿润。
低氧胁迫发芽:将发芽2 d后的苦荞放置于带盖的培养瓶(φ5 cm×18.5 cm)中,加入pH为4.2的10 mmol/L柠檬酸-柠檬酸钠缓冲溶液,培养液中通入速率为1.0 L/min的空气,通气量由空气流量计控制,然后置于培养箱中于31.3℃下继续培养2 d,培养期间,每隔24 h更换1次培养液。
培养结束后,将发芽种子用去离子水清洗并吸干表面水分,鲜样用于测定样品芽长和GAD活力,其余样品用真空冷冻干燥后去壳,粉碎至过80目筛后用于测定GABA含量。
1.3.2 低氧联合盐胁迫发芽试验设计
在低氧胁迫试验中,分别向培养液中添加0、5、10、20、30和50 mmol/L的NaCl溶液,培养结束后测定发芽苦荞芽长、GAD活力和GABA含量。
使用上一步骤中所得的最佳NaCl浓度,并在此基础上向培养液中分别添加0、1、3、5、7和9 mmol/L Ca2+(以CaCl2代替)溶液,研究低氧联合NaCl胁迫下Ca2+对发芽苦荞芽长、GAD活力和GABA含量的影响。
利用所得的最佳NaCl浓度和Ca2+溶液浓度,设计如下6个处理。正常发芽(对照):在培养皿中正常发芽4 d;低氧胁迫;低氧胁迫+10 mmol/L的NaCl;低氧胁迫+10 mmol/L的NaCl+3 mmol/L Ca2+;低氧胁迫+10 mmol/L的NaCl+3 mmol/L的钙离子螯合剂(EGTA);低氧胁迫+10 mmol/L的NaCl+ 3 mmol/L钙离子抑制剂(LaCl3),培养结束后测定发芽苦荞芽长、GAD活力和GABA含量。
1.3.3 测定指标与方法
芽长:随机选取30粒发芽苦荞籽粒,用游标卡尺测定其芽长;GABA:采用高压液相色谱HPLC,条件参照Bai等[23]的方法;GAD:参照Guo等[9]的方法。
1.4 数据处理与统计分析
试验设3次重复,结果以“¯x±s”表示。方差分析采用SPSS(version 16.0,Inc.,Chicago,IL,USA)软件,P<0.05表示差异显著,P<0.01表示差异极显著,不同小写字母之间表示差异显著(P<0.05)。
2 结果与分析
2.1 低氧胁迫下NaCl对发芽苦荞芽长、GAD活力和GABA富集量的影响
由表1可知,低氧胁迫下,用NaCl处理发芽苦荞,其芽长受到明显的抑制,当NaCl浓度超过5 mmol/L时,芽长显著下降(P<0.05)。发芽苦荞中GAD和GABA的含量随着NaCl浓度的增大均呈先上升后下降的趋势,并在10 mmol/L时达到最高值,分别为81.65 U/g FW和424.31 μg/g DW,分别是0 mmol/L的1.65倍和1.10倍。低氧联合10 mol/L的NaCl胁迫较单纯使用低氧胁迫相比可显著提高GABA的富集量(P<0.05)。
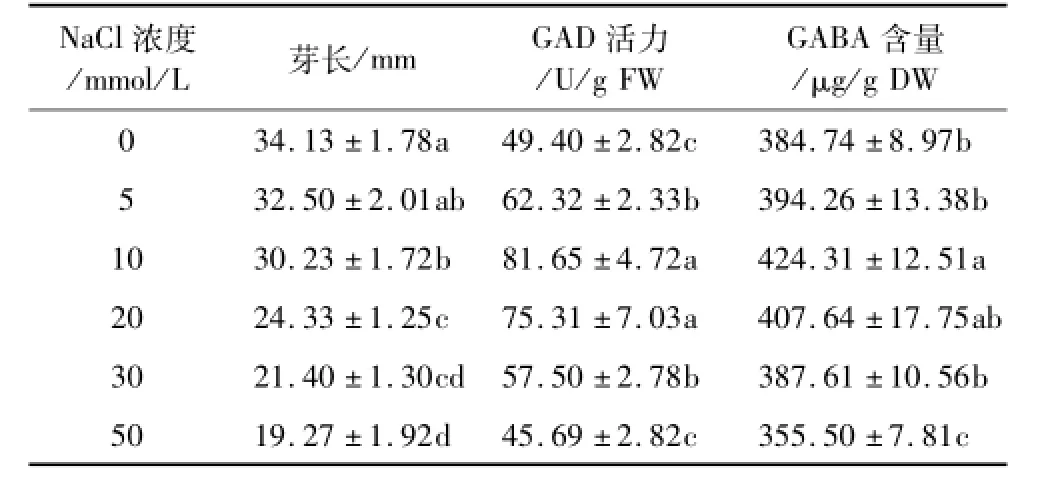
表1 NaCl对低氧胁迫下发芽苦荞芽长、GAD活力和GABA富集量的影响
2.2 低氧联合NaCl胁迫下Ca2+对发芽苦荞芽长、GAD活力和GABA富集量的影响
在低氧胁迫联合10 mmol/L的NaCl胁迫下,添加不同浓度的Ca2+,发芽苦荞芽长、GAD活性和GABA含量均发生了显著的变化(P<0.05)(见表2)。随着Ca2+浓度的提升,芽长不断增长,在3 mmol/L时达到最大为34.37 mm,较0 mmol/L显著提高(P<0.05);当Ca2+浓度超过3 mmol/L时,芽长极显著下降(P<0.01),苦荞芽长受到明显的抑制。GAD活力和GABA富集量随着Ca2+浓度的增高均呈现先升高后降低的趋势,在3 mmol/L Ca2+处理时达到最大,分别为91.44 U/g FW和492.31 μg/g DW,分别是0 mmol/L的1.14倍和1.16倍。适度浓度的Ca2+能够缓解NaCl的抑制作用,促进GAD活力的提高,进而使GABA富集量增大。
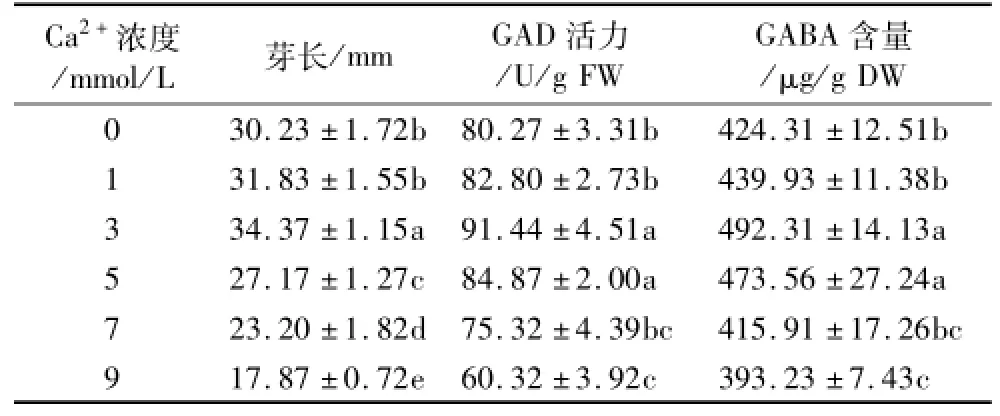
表2 Ca2+处理对发芽苦荞芽长、GAD活力和GABA富集量的影响
2.3 低氧联合NaCl胁迫下添加EGTA和LaC13对发芽苦荞芽长的影响
如图1所示,低氧胁迫抑制了苦荞的生长,其芽长为对照(正常培养)的77.24%,低氧胁迫下添加NaCl处理进一步抑制了苦荞芽长,此条件下芽长较单纯低氧胁迫减少了11.43%,在此条件下,添加适量的Ca2+处理使这种抑制作用得到缓解,与低氧联合NaCl胁迫相比芽长显著增加(P<0.05),但低于对照。在低氧联合NaCl胁迫基础上分别添加Ca2+的螯合剂EGTA和通道阻断剂LaC13处理后,芽长均显著降低(P<0.05),仅为24.30与25.33 mm,分别比低氧联合NaCl胁迫处理下降了19.63%和16.21%,二者处理之间无显著差异(P<0.05)。
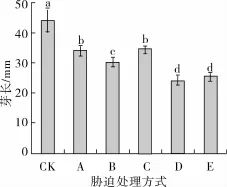
图1 不同胁迫方式对发芽苦荞芽长的影响
2.4 低氧联合NaCl胁迫下添加EGTA和LaC13对发芽苦荞GAD活力的影响
如图2所示,低氧胁迫下发芽苦荞GAD的活力较对照显著提升(P<0.05),低氧联合NaCl胁迫处理后,GAD活力极显著提升(P<0.01)。继续添加Ca2+处理,GAD活力大幅提升,可达91.44 U/g,较低氧联合NaCl胁迫提高了11.98%;而添加EGTA或LaC13处理后,GAD活力均会受到显著地抑制(P<0.05),仅分别为64.34和61.78 U/g FW,较低氧联合NaCl胁迫处理分别下降了21.20%和24.34%,说明Ca2+对GAD活力具有激活作用。
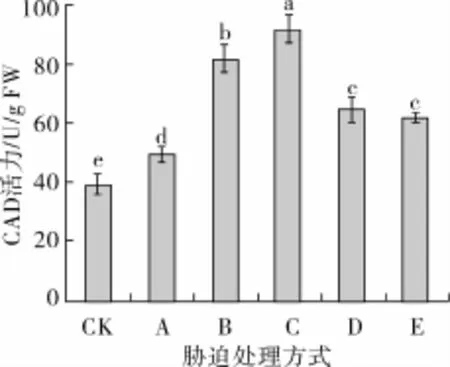
图2 不同胁迫方式对发芽苦荞GAD活力的影响
2.5 低氧联合NaCl胁迫下添加EGTA和LaC13对发芽苦荞GABA富集量的影响
不同胁迫处理对发芽苦荞GABA富集量的影响如图3所示。正常发芽4 d(对照)时苦荞GABA含量为187.39 μg/g DW,低氧胁迫后GABA含量较对照提升了1.05倍,为384.74 μg/g DW,继续联合NaCl处理,GABA含量增至424.31 μg/g DW,比低氧胁迫提升了10.29%,在此基础上,添加3 mmol/L的Ca2+后,GABA富集量可继续提升,达到492.32 μg/g DW,分别是低氧联合NaCl胁迫和对照的1.16和2.63倍。在低氧联合NaCl胁迫条件下,分别添加EGTA和LaC13处理均会显著降低GABA的富集量(P<0.05),仅分别为405.97和400.90 μg/g DW,二者处理间差异不显著(P<0.05)。
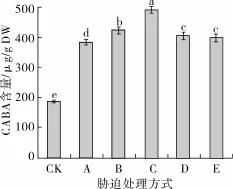
图3 不同胁迫方式对发芽苦荞GABA富集量的影响
3 讨论
植物在NaCl胁迫下的生长受到抑制[24]。NaCl胁迫损害植物细胞正常的代谢过程,最终导致植物生长不良[25],NaCl胁迫下大豆芽长较正常培养降低了12.28%~34.21%[26],发芽苦荞在低氧胁迫下添加NaCl后,其芽长明显下降。有研究报道,Ca2+能在一定程度上缓解这种胁迫作用,低浓度的Ca2+可增强淀粉酶活力[27],对糙米[27]NaCl胁迫下的大豆[28]和黄瓜种子[29]的萌发有促进作用,但高浓度的Ca2+则会出现抑制作用[29]。NaCl胁迫下,Ca2+通过增强信号转换和能量传输途径、促进蛋白质的生物合成、抑制蛋白质水解、对贮藏蛋白重新分配、调节内质网蛋白加工、激活并增强抗氧化酶的活性、积累次生代谢和渗透调节物质以及其他的适应性反应等方式来提高植物的耐盐性[28]。本研究在低氧联合NaCl胁迫的基础上添加3 mmol/L的Ca2+处理后,苦荞芽长显著大于单纯的低氧联合NaCl胁迫,而用5 mmol/L及以上的Ca2+处理则使芽长显著下降,表明较低浓度的Ca2+可改善低氧联合NaCl胁迫对苦荞生长的抑制,有促进苦荞生长的作用。
植物中的GABA合成主要来自GABA支路中由GAD催化的不可逆的α-谷氨酸脱羧反应,GAD是其限制酶,同时,该反应需要H+的参与[13]。植物受到低氧和涝灾、机械刺激、冷害、热激及盐胁迫等胁迫时会使细胞中的H+和Ca2+浓度增加强烈刺激GAD,导致体内GABA含量成倍增加[8,14-15],其中低氧胁迫下富集GABA最为快速有效,将植物放置于由通气的培养液形成的低氧环境中培养,不失为一种有效的低氧胁迫方法。Guo等[30]、Bai等[23]和Li等[31]分别将发芽大豆、粟谷和蚕豆置于通气的培养液中低氧培养,GAD活力大幅提升,GABA富集量得到快速增加。将发芽苦荞置于通气速率为1.0 L/min的条件下进行低氧胁迫,较正常培养相比,GABA富集量得到显著提升,与前述研究相一致。GAD是一种Ca2+/CaM依赖型酶,具有一个钙调蛋白结合区,增加细胞内Ca2+浓度,可促进CaM转录水平提高,CaM调控的一系列生理反应随之被激活,继而提升GAD活性[19]。研究表明,NaCl胁迫可通过提高细胞内Ca2+浓度来刺激GAD活性,从而增加GABA含量。陈惠等[32]发现0~40 mmol/L的NaCl处理提高了发芽蚕豆中GAD活性,进而使GABA的含量升高。NaCl胁迫诱导紫花苜蓿根中氨基酸和碳水化合物大量合成,GABA含量亦有所提升[33]。在一定范围内,NaCl胁迫下发芽大豆[34]和玉米幼苗中[35]GABA的富集量随着胁迫强度的增大而提升。低氧胁迫引起GABA含量的提高被认为是由于H+浓度的增加继而刺激GAD活性所导致[36],本研究中,低氧联合NaCl处理显著提高了发芽苦荞GAD活力,较单纯低氧胁迫提高了65.30%,GABA富集量得到迅速提升。低氧联合NaCl胁迫对GAD的提升和GABA的富集有叠加的效应,比单纯使用低氧和NaCl胁迫要高。
研究表明,添加外源Ca2+处理可促进发芽糙米[37]和蚕豆[38]的GAD活力,进而富集GABA。Guo等[9]在低氧胁迫下对发芽大豆分别使用Na+和Ca2+处理,诱导了GAD活力的提升,提升了GABA的含量。在NaCl胁迫的基础上,若继续添加Ca2+处理,可提升GABA的富集效果。Bai等[19]报道,NaCl和Ca2+联合胁迫发芽粟谷可使其GAD活力提升,GABA富集量最高可较单纯NaCl胁迫增加36.29%。Ca2+不仅可以激活GAD的活性,还能促进相关基因的表达[18]。Yin等[15]报道,外源Ca2+可显著增强NaCl胁迫下发芽大豆GAD相关基因的表达,促进GAD活力的提升。本研究表明,低氧联合NaCl胁迫下,添加一定浓度的Ca2+同样可使GAD活力大幅提升,GABA的富集量达到最大,是低氧联合NaCl胁迫的1.16倍,是对照的2.63倍。EGTA和LaC13分别是Ca2+的螯合剂和通道阻断剂,前者通过对Ca2+进行螯合继而消除Ca2+的生理作用,后者阻碍了质膜上的钙通道,且浓度较高时完全抑制Ca2+通过质膜进入细胞以达到对植物细胞生长的负面作用[24]。采用EGTA和LaC13处理过的发芽粟谷[19]和发芽蚕豆[39]中芽长、GAD活力和GABA含量较对照显著降低。本研究表明,低氧联合NaCl胁迫处理下,3 mmol/L Ca2+处理可缓解NaCl胁迫对苦荞芽长的抑制作用,同时提高了GAD的活性,进而使苦荞芽中GABA的含量大幅度提升;而添加EGTA和LaC13可抑制Ca2+/CaM的形成,阻断钙信号的发生,从而降低GAD活力,使GABA的富集量下降。研究进一步证实了GAD是一种由Ca2+/CaM调节的酶,但低氧联合NaCl胁迫下Ca2+与GAD活力及GABA的富集的作用机理有待深入研究。
4 结论
低氧联合NaCl胁迫对苦荞的芽长有抑制作用,GAD活力和GABA的含量随着NaCl浓度的增大均呈先上升后下降的趋势,在10 mmol/L NaCl溶液处理时均达到最高值,分别是单纯低氧胁迫的1.65和1.10倍;在低氧联合NaCl胁迫的基础上添加低浓度的Ca2+可缓解NaCl胁迫对苦荞芽长的抑制作用,添加3 mmol/L Ca2+处理可使GAD活力和GABA的富集量达到最大,分别是低氧联合NaCl胁迫的1.12倍(91.44 U/g FW)和1.16倍(492.32 μg/g DW);低氧联合NaCl胁迫下添加Ca2+的螯合剂或通道阻断剂后,发芽苦荞芽长显著降低,GAD活力显著下降,GABA富集量减少。
[1]Hong Y H,Chao W W,Chen M L,et al.Ethyl acetate extracts of alfalfa(Medicago sativa L.)sprouts inhibit lipopolysaccharide-induced inflammation in vitro and in vivo[J]. Journal of Biomedical Science,2009,16:64-64
[2]Martinez-Villaluenga C,Kuo Y H,Lambein F,et al.Kinetics of free protein amino acids,free non-protein amino acids and trigonelline in soybean(Glycine max L.)and lupin(Lupinus angustifolius L.)sprouts[J].European Food Research and Technology,2006,224(2):177-186
[3]Kim S L,Kim S K,Park C H.Introduction and nutritional evaluation of buckwheat sprouts as a new vegetable[J].Food Research International,2004,37(4):319-327
[4]Kitazaki K,Watanabe S,Okamoto A,et al.Far-red light enhances removal of pericarps in tartary buckwheat(Fagopyrum tataricum Gaertn.)sprout production under artificial lighting[J].Scientia Horticulturae,2015,185:167-174
[5]陈尚钘,王宗德,陈宏伟,等.添加锌素营养液对培养苦荞芽菜品质的影响[J].中国食品学报,2006,6(6):47-51 Chen S Y,Wang Z D,Chen H W,et al.The effect of Znadded nutrient fluid on the quality of tratary buckwheat bud[J].Journal of Chinese Institute of Food Science and Technology,2006,6(6):47-51
[6]Huang X Y,Zeller F J,Huang K F,et al.Variation of major minerals and trace elements in seeds of tartary buckwheat(Fagopyrum tataricum Gaertn.)[J].Genetic Resources and Crop Evolution,2014,61(3):567-577
[7]Li D,Li X L,Ding X L.Composition and Antioxidative properties of the flavonoid-rich fractions from tartary buckwheat grains[J].Food Science and Biotechnology,2010,19(3):711-716
[8]Bouche N,Lacombe B,Fromm H.GABA signaling:a conserved and ubiquitous mechanism[J].Trends in Cell Biology,2003,13(12):607-610
[9]Guo Y X,Yang R Q,Chen H,et al.Accumulation of γaminobutyric acid in germinated soybean(Glycine max L.)in relation to glutamate decarboxylase and diamine oxidase activity induced by additives under hypoxia[J].European Food Research and Technology,2012,234(4):679-687
[10]朱云辉,郭元新.发芽苦荞GABA的富集及氨基酸等含氮物的变化[J].食品与发酵工业,2015,41(2):85-88 Zhu Y H,Guo Y X.Study on the accumulation and changes of γ-aminobutyric acid,amino acid and other nitrogen compounds in germinating tartary buckwheat[J].Food and Fermentation Industries,2015,41(2):85-88
[11]Abdou A M,Higashiguchi S,Horie K,et al.Relaxation and immunity enhancement effects of γ-Aminobutyric acid(GABA)administration in humans[J].Biofactors,2006,26(3):201-208
[12]Vidal-Valverde C,Frias J,Sierra I,et al.New functional legume foods by germination:effect on the nutritive value of beans,lentils and peas[J].European Food Research and Technology,2002,215(6):472-477
[13]Shelp B J,Bown A W,McLean M D.Metabolism and functions of gamma-aminobutyric acid[J].Trends in Plant Science,1999,4(11):446-452
[14]Kinnersley A M,Turano F J.Gamma aminobutyric acid(GABA)and plant responses to stress[J].Critical Reviews in Plant Sciences,2000,19(6):479-509
[15]Yin Y Q,Yang R Q,Guo Q H,et al.NaCl stress and supplemental CaCl regulating GABA metabolism pathways in germinating soybean[J].European Food Research&Technology,2014,238(5):781-788
[16]Chung H J,Jang S H,Cho H Y,et al.Effects of steeping and anaerobic treatment on GABA(gamma-aminobutyric acid)content in germinated waxy hull-less barley[J]. LWT-Food Science and Technology,2009,42(10):1712-1716
[17]Reggiani R,Cantu C A,Brambilla I,et al.Accumulation and interconversion of amino acids in rice roots under anoxia[J].Plant Cell Physiology,1988,29:981-987
[18]Yang R Q,Guo Y X Wang S F,et al.Ca2+and aminoguanidine on γ-aminobutyric acid accumulation in germinating soybean under hypoxia-NaCl stress[J].Journal of Food and Drug Analysis,2015,23(2):287-293
[19]Bai Q Y,Yang R Q,Zhang L X,et al.Salt stress induces accumulation of γ-aminobutyric acid in germinated foxtail millet(Setaria italica L.)[J].Cereal Chemistry,2013,90(2):145-149
[20]韩永斌,顾振新,蒋振辉.Ca2+浸泡处理对发芽糙米生理指标和GABA等物质含量的影响[J].食品科学,2006,27(10):58-61 Han Y B,Gu Z X,Jiang Z H.Effect of Ca2+on some physiological indexes and contents of γ-aminobutyric acid and other essential substances in germinating brown rice immersed into aerobic water[J].Food Science,2006,27(10):58-61
[21]Peng L X,Zou L,Zhao J L,et al.Response surface modeling and optimization of ultrasound-assisted extraction of three fla-vonoids from tartary buckwheat(Fagopyrum tataricum)[J]. Pharmacognosy Magazine,2013,9(35):210-215
[22]Zielinska D,Turemko M,Kwiatkowski J,et al.Evaluation of flavonoid contents and antioxidant capacity of the aerial parts of common and tartary buckwheat plants[J].Molecules,2012,17(8):9668-9682
[23]Bai Q Y,Fan G J,Gu Z X,et al.Effects of culture conditions on gamma-aminobutyric acid accumulation during germination of foxtail millet(Setaria italica L.)[J].European Food Research and Technology,2008,228(2):169-175
[24]白青云.低氧胁迫和盐胁迫下发芽粟谷γ-氨基丁酸富集机理及抗氧化性研究[D].南京:南京农业大学,2009 Bai Q Y.Studies on mechanism of γ-aminobutyric acid accumulation and antioxidant activity in germinated foxtail millet under hypoxia stress and salt stress[D].Nanjing:Nanjing Agricultural University,2009
[25]尚庆茂,宋士清,张志刚,等.水杨酸增强黄瓜幼苗耐盐性的生理机制[J].中国农业科学,2007,40(1):147-152 Shan Q M,Song S Q,Zhang Z G,et al.Physiological mechanisms of salicylic acid enhancing the salt tolerance of cucumber seedling[J].Scientia Agricultura Sinica,2007,40(1):147-152
[26]郭元新.盐和低氧胁迫下发芽大豆γ-氨基丁酸富集与调控机理研究[D].南京:南京农业大学,2011 Guo Y X.Accumulation of γ-aminobutyric acid and its regulation mechanisms under NaCl and hypoxia stress in germinated soybean[D].Nanjing:Nanjing Agricultural U-niversity,2011
[27]郑艺梅.发芽糙米营养特性、γ-氨基丁酸富集及生理功效的研究[D].武汉:华中农业大学,2006 Zheng Y M.Study of germinated brown rice on its nutritional property and physiological function of gamma aminobutyric acid accumulation therein[D].Wuhan:Huazhong Agricultural University,2006
[28]Yin Y Q,Yang R Q,Han Y B,et al.Comparative proteomic and physiological analyses reveal the protective effect of exogenous calcium on the germinating soybean response to salt stress[J].Journal of Proteomics,2015,113:110-126
[29]王广印,张百俊,赵一鹏,等.NaCl胁迫对黄瓜种子萌发的影响[J].吉林农业大学学报,2004,26(6):624-627 Wang G Y,Zhang B J,Zhao Y P,et al.Effects of NaCl stress on seed germination of cucumber[J].Journal of Jilin Agricultural University,2004,26(6):624-627
[30]Guo Y X,Chen H,Song Y,et al.Effects of soaking and aeration treatment on γ-aminobutyric acid accumulation in germinated soybean(Glycine max L.)[J].European Food Research and Technology,2011,232(5):787-795
[31]Li Y,Bai Q Y,Jin X J,et al.Effects of cultivar and culture conditions on γ-aminobutyric acid accumulation in germinated fava beans(Vicia faba L.)[J].Journal of the Science of Food and Agriculture,2010,90(1):52-57
[32]陈惠,杨润强,李岩,等.氯化盐和激素对发芽蚕豆中γ-氨基丁酸富集的影响[J].南京农业大学学报,2012,35(1):119-124 Chen H,Yang R Q,Li Y,et al.Effects of chloride salts and hormones on γ-aminobutyric acid enrichment in germinated faba beans(Vicia faba L.)[J].Journal of Nangjing Agricultural University,2012,35(1):119-124
[33]Fougère F,Le Rudulier D,Streeter J G.Effects of salt stress on amino acid,organic acid,and carbohydrate composition of roots,bacteroids,and cytosol of alfalfa(Medicago sativa L.)[J].Plant Physiology,1991,96(4):1228-1236
[34]郭元新,杨润强,陈惠,等.盐胁迫富集发芽大豆γ-氨基丁酸的工艺优化[J].食品科学,2012,33(10):1-5 Guo Y X,Yang R Q,Chen H,et al.Optimization of GABA accumulation process of germinated soybean under salt stress[J].Food Science,2012,33(10):1-5
[35]周翔,吴晓岚,李云,等.盐胁迫下玉米幼苗ABA和GABA的积累及其相互关系[J].应用与环境生物学报,2005,11(4):412-415 Zhou X,Wu X L,Li Y,et al.Accumulations and correlations of ABA and GABA in maize seedling under salt stress[J].Chinese Journal of Applied and Environmental Biology,2005,11(4):412-415
[36]Scott-Taggart C P,Van Cauwenberghe O R,McLean M D,et al.Regulation of gamma-aminobutyric acid synthesis in situ by glutamate availability[J].Physiologia Plantarum,1999,106(4):363-369
[37]肖君荣.发芽处理对糙米中GABA含量影响及其蒸煮食用品质研究[D].广州:华南理工大学,2014 Xiao J R.Study on effectsof germination on thecontent of GABA and edible quality of brown rice[D].Guangzhou:South China University of Technology,2014
[38]陈惠,杨润强,韩永斌,等.发芽蚕豆富集γ-氨基丁酸的培养液组分优化[J].中国粮油学报,2011,26(11):27-31 Chen H,Yang R Q,Han Y B,et al.Optimization of culture solution compositions for γ-aminobutyric acid accumulation germinated fava beans(Vicia faba L.)[J].Journal of the Chinese Cereals and Oils Association,2011,26(11):27-31
[39]陈惠.发芽蚕豆γ-氨基丁酸富集与调控技术研究[D].南京:南京农业大学,2012 Chen H.Study on γ-aminobutyric acid accumulation and regulation techniques in germinated fava benas(Vicia faba L.)[D].Nanjing:Nanjing Agricultural University,2012.
Effect of Exogenous Ca2+on GABA Accumulation in Germinated Tartary Buckwheat Under Hypoxia-NaCl
Zhu Yunhui Guo Yuanxin Du Chuanlai Ding Zhigang Ye Hua
(College of Food and Drug,Anhui Science and Technology University,Fengyang 233100)
The effect of exogenous Ca2+on GABA accumulation in germinated tartary buckwheat under hypoxia-NaCl stress was investigated by adding exogenous Ca2+to the aeration medium of the aeration medium.The tartary buckwheat sprouts length was restrained under hypoxia-NaCl stress in which GAD activity and GABA content rise at first and then drop with the increase of salt concentration.The maximum were reached at the 10 mmol/L salt solution,and were 1.65 times higher and 1.10 times higher than that under hypoxia stress alone respectively.On this basis,an addition of a low concentration of Ca2+in hypoxia-NaCl stress could alleviate salt stress inhibition of the tartary buckwheat sprouts length.The addition of 3 mmol/L Ca2+could make the GAD activity and GABA concentration reach the maximum,which were 91.44 U/g FW and 492.31 μg/g DW,1.12 times higher and 1.16 times higher than that under hypoxia-NaCl stress respectively.Under hypoxia-NaCl stress,the germinated tartary buckwheat sprouts length decreased obviously,GAD activity was significantly decreased and GABA concentration decreased after the addition of Ca2+chelating agent and channel blockers.Exogenous Ca2+could relieve the salt stress on the tartary buckwheat sprouts length inhibition and enhanced GABA accumulation in germinated tartary buckwheat through the improvement of GAD activity.
hypoxia stress,NaCl stress,Ca2+treatment,germinated tartary buckwheat,GABA accumulation
TS201.1
A
1003-0174(2017)01-0017-07
安徽省自然科学基金(1308085MC32)
2015-06-13
朱云辉,男,1991年出生,硕士,食品功能性成分的富集及品质控制
郭元新,男,1970年出生,教授,农产品加工及品质控制
猜你喜欢
农业科技与信息(2021年8期)2021-12-06
诗潮(2021年11期)2021-11-24
中国食品(2021年17期)2021-09-27
四川农业与农机(2021年1期)2021-01-05
浙江农业科学(2020年10期)2020-11-06
现代园艺(2020年7期)2020-04-22
中国动物传染病学报(2020年2期)2020-04-02
湖北农业科学(2019年20期)2019-12-11
中国粮油学报(2019年4期)2019-07-12
科学与财富(2018年8期)2018-05-09
