
Cd2+、Cu2+、Zn2+、Pb2+对菹草生长胁迫影响及其应答机制
2016-11-10 09:28李婉璐张光生成小英
食品与生物技术学报 2016年9期
李婉璐, 张光生, 成小英*
(1.江南大学 环境与土木工程学院 江苏 无锡 214122;2.江苏省水处理技术与材料协同创新中心,江苏 苏州215000)
Cd2+、Cu2+、Zn2+、Pb2+对菹草生长胁迫影响及其应答机制
李婉璐1,2,张光生1,2,成小英*1,2
(1.江南大学 环境与土木工程学院江苏 无锡 214122;2.江苏省水处理技术与材料协同创新中心,江苏 苏州215000)
为探求不同浓度的Cd2+、Cu2+、Zn2+、Pb2+对菹草(石芽)的生长发育的胁迫及其应答机制,对菹草(石芽)的生长状况、叶片的叶绿素、可溶性蛋白质、丙二醛(MDA)含量、抗氧化酶系统等进行了分析。结果如下:Cu2+、Cd2+质量浓度大于0.5 mg/L,植株无法正常生长或存活;Cu2+胁迫下,植株受损程度相对严重。Zn2+、Pb2+质量浓度小于10mg/L时,菹草仍可以正常生长,但Pb2+胁迫程度比Zn2+较轻。综上,在相同质量浓度下,4种重金属对菹草(石芽)的胁迫强弱依次为Cu2+、Cd2+、Zn2+、Pb2+。本研究为生态修复中水生植物的金属阈值提供了有价值的参考。
胁迫;菹草;Cd2+;Cu2+;Zn2+;Pb2+
目前,水生植物作为水体主要的初级生产者,为水生生态系统结构和功能的维持起着不可替代的作用[1-2]。一些沉水植物如马来眼子菜、苦草、菹草等,因其对水体营养物质和重金属的吸收富集作用以及对浮游藻类的竞争抑制,已成为水体原位富营养化控制和重金属治理的一项重要生物措施[3-4]。然而,过高浓度的营养盐,特别是高浓度的金属元素,又会反过来对水生植物的生长发育产生胁迫[5]。因此,有必要就重金属离子对水生植物的发育及生长的生理生化影响进行研究。
近年来,国内外学者进行了一些关于重金属胁迫水生植物的研究,李玥[5]以镉、铜、锌对4种水生植物的毒害分析其应答机制,陈辉[6]等通过Cr6+对菹草的胁迫过程中叶绿素荧光参数,抗氧化系统和超微结构的变化探究其应答机制,Uruc[3]等研究了3种浮萍在Zn2+胁迫下抗氧化系统的响应。但这些研究多是采用成体进行实验,而忽视了重金属胁迫对菹草从石芽幼体到成体的过程影响。
作者选取菹草石芽为培养对象,研究了在菹草一个生长周期内,4种重金属离子胁迫的浓度效应与菹草抗氧化酶体系的应答机制,为污染水体中菹草无性生殖生长周期内的重金属胁迫与耐受性机制提供理论依据。
1 材料与方法
1.1材料与处理
试验所用菹草于2013年11月采自江南大学小蠡湖水域。取水体底泥中未发芽的菹草石芽,先以清水洗净,再依次用5%的H2O2(15 min)、10%的NaClO浸泡消毒30 s,并用无菌水清洗6遍[7]。然后将这些石芽置于重金属处理的1/10 Hoagland灭菌培养液中,在光照培养箱中培养,光照周期为12 h∶12 h(光∶暗),光照强度为8 000 Lux,光暗温度为25℃∶18℃(光∶暗)。每3天更换培养液,直至石芽发育生长为成熟的菹草并长出新的石芽 (约1个月)后停止培养。本试验共分为25组:对照组(CK)用1/10 Hoaglands营养液培养,其余24组分别用添加了0.1、0.5、1.0、2.5、5.0、10 mg/LZn2+(ZnSO4·7H2O)、Cd2+(CdSO4)、Cu2+(CuSO4·5H2O)或Pb2+(Pb(NO3)2)的1/10 Hoaglands营养液培养,每组设3个重复,每个重复20颗石芽。
1.2测定方法
每日统计单颗菹草石芽的萌发芽数、生根数并计算每组实验处理的萌发率。叶绿素含量采用二甲亚砜法测定[8]。可溶性蛋白质含量采用Bradford的考马斯亮蓝G-250法测定[9]。MDA含量采用硫代巴比妥酸法(TBA法)测定[9]。超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)的含量采用植物elisa检测试剂盒(上海颖心实验室设备有限公司)测定。
1.3统计分析
所有数据均为3次独立实验的平均值±标准差(Mean±SD),并用SPSS 19.0进行方差分析,经分析有效的数据(p<0.05)进行LSD多重分析比较。
2 结果与分析
2.1Cd2+、Cu2+、Zn2+、Pb2+对石芽发芽率和生根率的影响
实验处理5 d后,各组石芽发芽率均达到100%,单颗石芽的出芽数在1~5之间,不同金属及不同金属浓度处理下的出芽数无显著差异。除Cu2+质量浓度为10mg/L时的生根率在50%,其余各组石芽的生根率均在90%以上。各组的单颗石芽平均生根数有明显变化,见图1。Zn2+和Pb2+胁迫组中,平均生根数没有随金属浓度的增加而降低;而Cd2+组中生根数自质量浓度大于1mg/L有微幅下降;Cu2+胁迫组在质量浓度等于10mg/L时生根数明显低于其他胁迫组。
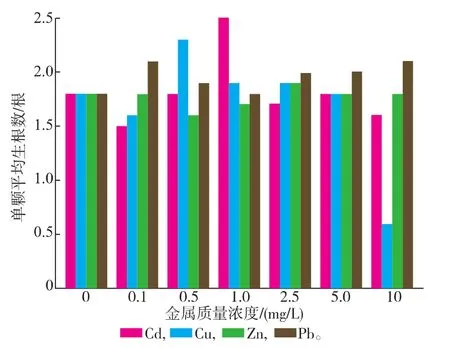
图1 Cd2+、Cu2+、Zn2+、Pb2+对石芽生根数的影响Fig.1 Effects of Cd2+,Cu2+,Zn2+,Pb2+stress on the numbers of w inter buds,Potamogeton crispus L.
不同质量浓度的Cd2+、Cu2+、Zn2+、Pb2+对菹草生长发育的影响见表1。Zn2+和Pb2+胁迫组中,石芽逐渐发育为成熟植株,且长势较好,与对照组无显著差异;而Cu2+胁迫组中,当质量浓度≥1 mg/L时植株叶片出现枯黄和暗斑,质量浓度在5mg/L时植株矮小,叶片全部枯黄,质量浓度在10mg/L时植物仍然处于石芽出芽状态,未生叶,无法进行后续检测,同时培养液易浑浊;Cd2+胁迫组中,质量浓度在2.5 mg/L时植株已经褪绿,5.0 mg/L时植物整体出现萎蔫现象,在10.0mg/L质量浓度下植株矮小,根系脱落,叶片细小且枯萎,无法进行后续检测。
综合Cd2+、Cu2+、Zn2+、Pb2+胁迫对菹草生长发育情况的表观影响,各金属离子对菹草损伤程度为:Cu2+>Cd2+>Pb2+、Zn2+。

表1 Cd2+、Cu2+、Zn2+、Pb2+胁迫下植株生长情况Table 1 Grow th state of plants under stress of Cd2+,Cu2+,Zn2+,Pb2+
2.2Cd2+、Cu2+、Zn2+、Pb2+胁迫对菹草叶绿素、可溶性蛋白质质量分数的影响
与对照组相比,Cd2+和Cu2+各质量浓度处理均明显降低了菹草叶片叶绿素质量分数,而Zn2+和Pb2+胁迫组各浓度处理对菹草叶片叶绿素质量分数的影响并不显著,见图2。同质量浓度Cd2+和Cu2+胁迫下,Cd2+组的菹草叶绿素质量分数均高于Cu2+组。

图2 Cd2+、Cu2+、Zn2+、Pb2+胁迫对菹草叶片叶绿素、可溶性蛋白质质量分数的影响Fig.2 Effects of Cd2+,Cu2+,Zn2+,Pb2+stress on the contents of chlorophyll and soluble protein in the leaves of Potamogetous crispus L.
在4种金属不同质量浓度梯度胁迫下,菹草可溶性蛋白质质量分数与对照组相比均表现为先上升后下降。当Cd2+或Cu2+的质量浓度≤0.1mg/L时,可溶性蛋白质平缓上升;而当Cd2+或Cu2+≥0.5mg/L时,可溶性蛋白质开始显著下降;当质量浓度为10 mg/L时,由于植株未出叶(Cu2+胁迫组)或完全腐烂(Cd2+胁迫组),无法取样测试。在Zn2+胁迫下,在质量浓度≤5.0mg/L时,随着金属质量浓度的增加,可溶性蛋白质质量分数呈先上升后下降的趋势,且均高于对照组,而质量浓度为10 mg/L时,质量分数显著下降,低于对照组;而Pb2+胁迫组中,以0.5 mg/L处理组为分界线,可溶性蛋白质质量分数先上升,后平缓下降。由此可认为,各金属离子对菹草可溶性蛋白质质量分数的影响为:Cu2+>Cd2+>Pb2+>Zn2+。
2.3Cd2+、Cu2+、Zn2+、Pb2+胁迫对菹草SOD,POD,CAT活性和MDA摩尔质量浓度的影响
Cd2+胁迫下,抗氧化酶SOD、POD、CAT活性变化随质量浓度的升高先上升后下降,分别在0.5、0.1、0.5 mg/L时达到最大值。在5.0mg/L时3种酶活性仅为CK组的50%、30%、44%。MDA含量总体呈上升趋势,在5.0mg/L时达到最大值,见表2。
Cu2+胁迫下,质量浓度≤0.1 mg/L时,SOD、POD、CAT活性相比对照组微量上升,但低于相应的Cd2+处理组。随着处理质量浓度的继续上升,3种酶活性呈持续下降的趋势,均在5.0 mg/L浓度时降到底值,分别为对照组的63%、41%、42%。MDA摩尔质量浓度变化为平缓上升,在5.0 mg/L达到峰值。
Zn2+胁迫组下,随处理质量浓度的升高,SOD活性变化先上升后下降,在质量浓度0.5 mg/L达到最大值,增幅为20%;质量浓度5mg/L时,活性有所下降但仍高于对照组,10 mg/L时仅为对照组的89%。低质量浓度(≤1.0 mg/L)下,POD和CAT活性变化不显著,与对照组差异不大;高质量浓度下(≥2.5 mg/L),POD和CAT均呈下降趋势,且POD活性下降更迅速,两种酶均在10mg/L达到最小值,与对照相比降幅分别为63%和23%。MDA摩尔质量浓度先迅速上升后平缓下降,最小值(5 mg/L)仍高于对照组,增幅为30%。之后MDA摩尔质量浓度缓慢上升,最大值(10mg/L)高于对照组。
在Pb2+胁迫处理中,SOD、POD、CAT活性的变化趋势与Zn2+胁迫组相似,随处理质量浓度的增加先平缓上升后下降,均在0.5 mg/L达到最大值。MDA摩尔质量浓度在低质量浓度下变化显著,在高质量浓度(10mg/L)下,显著上升,增幅为61%。

表2 Cd2+、Cu2+、Zn2+、Pb2+胁迫对菹草叶片SOD,POD,CAT活性和MDA含量的影响Table 2 Effects of Cd2+,Cu2+,Zn2+,Pb2+stress on the contents of MDA and the activities of SOD,POD,CAT in the leaves of Potamogetous crispus L.
3 结语
本研究表明,石芽在Zn2+和Pb2+胁迫(≤10.0 mg/L)下,仍能正常完成一个生长周期,且植株生长良好。相比之下,Cd2+对植株的毒害作用较强,Cu2+则更甚之。
叶绿素质量分数是反映植物光合器官生理状况的重要指标。本研究结果显示,低质量浓度(≤0.1 mg/L)Cd2+和Cu2+处理后的菹草生长较为良好,与对照组差异不显著;但随着处理质量浓度的进一步升高,菹草褪绿现象极为明显,尤其是在高质量浓度(≥5 mg/L)处理下,叶片枯黄,带有死斑,与叶绿素质量分数变化相一致。在10mg/L时,Cd2+处理组叶片发黑,Cu2+组中菹草由于长时间胁迫,石芽未生叶。叶绿素的质量分数降低可能是由于高质量浓度重金属抑制或破坏了合成叶绿素所需的酶(如叶绿素酸酯还原酶),亦或是由于氨基-r-酮戊酸的合成受阻[10]。李玥等[5]的研究表明,Cu2+、Cd2+等重金属离子对叶绿素中心Mg原子(叶绿素组成成分之一)的取代会导致光合作用中断。而Zn2+各处理组中叶绿素质量分数(相对于CK组)差异并不显著,可能是由于Zn2+是叶绿体结构的必要组成成分[11],低质量浓度Zn2+能够促进叶绿素合成。且质量浓度到达10 mg/L时,对菹草的毒害作用并不显著,菹草仍能生长完好。与Zn2+胁迫相似,Pb2+各处理组菹草的叶绿素质量分数并没有显著减少。同质量浓度下,Cu2+处理组的叶绿素质量分数低于Cd2+处理组,可见Cu2+对菹草光合器官的毒害作用强于Cd2+;同时Cu2+处理组易产生Cu(OH)2沉淀,导致培养液浑浊,直接减弱光照强度,可能对植株生长也有一定的抑制作用,。
可溶性蛋白质质量分数的高低能反映植物生理活动的旺盛程度[12],4种金属胁迫组的可溶性蛋白质质量分数均随着质量浓度的升高不同程度地呈现先上升后下降的趋势。其中Cu2+对菹草的毒害作用最显著,导致可溶性蛋白质质量分数迅速降低,其次是Cd2+、Pb2+、Zn2+。这可能因为Cu2+在细胞内促进了蛋白水解酶的活性,加速了蛋白质的水解[13],也可能是使蛋白质合成的相关细胞器受到损伤,在DNA转录翻译、蛋白质合成的某一或某些环节产生毒害和抑制作用。Cd2+和Pb2+可能也是相似的毒害机理,而Zn2+作为植物生长必需元素,在质量浓度≤5.0mg/L时,可溶性蛋白质质量分数均高于CK组。由此推测,低质量浓度(≤5.0mg/L)的Zn2+能够参与植物体内代谢,促进蛋白质合成,但高质量浓度的Zn2+则会抑制蛋白质的合成,或是阻碍DNA转录翻译途径,从而影响蛋白质的合成[14]。
在逆境条件下,植物体内的活性氧代谢平衡被打破,产生大量活性氧物质,如O-2·和H2O2等,一定量的累积便会损伤细胞进而影响植物生长[15]。研究表明[16],O-2·的过量累积导致膜脂过氧化或膜脂脱脂作用产生MDA,从而破坏膜结构。因此,MDA摩尔质量浓度是反映膜脂过氧化作用的强弱和质膜受破坏程度的重要指标[17]。SOD、POD和CAT是植物抗氧化保护系统中的主要酶。其中SOD将O-2·歧化成H2O2,再由CAT、POD催化H2O2分解成H2O,从而缓解O-2·和H2O2的积累[15]。3种酶协同作用下有利于清除过多的活性氧并使之维持在较低水平[18]。姜岩[19]研究表明,低浓度Cu2+在短期胁迫下能够使SOD、POD和CAT活性逐渐增加,是植物对防止活性氧伤害的一种保护性适应;杨红玉[20]等研究表明,低质量浓度Cd2+可以促进植物呼吸酶和三羧酸循环,然而随着Cd2+质量浓度升高,3种酶活性受抑制,叶片内的抗氧化酶不足以抵御活性氧含量的升高,在高质量浓度下,菹草均表现出明显的叶损伤,叶绿素显著减少。
在本研究中,在高质量浓度Cd2+、Cu2+胁迫下,SOD虽起主要的保护作用,但由于在完成一个生长周期后,植株受到长达1个月的重金属胁迫,SOD活性也显著下降,低于CK组;同时POD和CAT的保护程度降低,POD、CAT也是活性氧清除系统的重要保护剂,它们活性的降低又在一定程度上加重了活性氧对膜的伤害[21],进而导致植物细胞受损过大,保护能力显著下降,细胞结构及生理生化反应受到破坏,最终加速植物的衰老和死亡[22]。
Zn2+和Pb2+胁迫组中,SOD、POD和CAT活性变化随处理浓度的升高呈一定程度的先上升后下降的趋势,在质量浓度为2.5mg/L时,SOD活性仍高于CK组,且最小值也为CK组的89%。在高质量浓度(≥5 mg/L)下,SOD起主要的保护作用,而POD和CAT的保护程度虽有降低,但相比于同浓度的Cd2+、Cu2+胁迫,Zn2+和Pb2+胁迫组的酶防御能力相对较强。而MDA摩尔质量浓度均高于CK组,可能是长时间低质量浓度(≤0.1 mg/L)的胁迫,导致植物产生大量过氧化物MDA,此时SOD活性正在提高,未及时清除MDA;随着处理质量浓度的升高,MDA摩尔质量浓度先下降再上升,说明SOD对植物膜系统保护作用是有一定限度的,高质量浓度Zn2+和Pb2+胁迫致使植株膜脂过氧化作用增强,但最高增幅分别仅为109%和61%,且SOD、POD、CAT活性仍较好,植株在外部形态上仍表现完好。
同质量浓度下,Cu2+胁迫对菹草的胁迫作用最显著,其次是Cd2+,最后是Pb2+和Zn2+。低质量浓度的Zn2+(≤5.0mg/L)对菹草的生长有一定的促进作用,但高质量浓度的Zn2+(10.0 mg/L)也会对植物产生毒害作用,即植物对超过需求量的Zn2+的胁迫也非常敏感。低质量浓度Pb2+(≤10.0mg/L)胁迫对菹草的毒害作用不显著,外部形态完好,但是SOD、POD、CAT活性及可溶性蛋白质质量分数变化显著,后期会出现叶片褪绿等现象。Cd2+、Cu2+胁迫对石芽发育有明显的毒害作用,少量便可抑制菹草叶绿素和可溶性蛋白质的合成,更高质量浓度下,植株直接停止生长。
4种金属均不同程度地加快了MDA的产生。随着处理质量浓度的升高,SOD、POD、CAT活性先上升后下降,表明植物在受到超需求的金属胁迫-下,会通过自身的抗氧化系统及时清除H2O2和O2·等过多活性氧来增强抗逆性;但当重金属浓度超过植物的耐受力后,植物的抗氧化系统不足以抵御不断产生的大量活性氧,且合成受到抑制,植物的叶绿素和可溶性蛋白质显著减少,表现出明显的损伤现象。
[1]焦伟,卢少勇,李光德,等.环太湖主要进出河流重金属污染及其生态风险评价[J].应用与环境生物学报,2010,16(4):577-580. JIAOWei,LU Shaoyong,LIGuangde,etal.Heavymetalpollution ofmain inflow and outflow rivers around the Taihu lake and assessment of its potential ecological risk[J].Journal of Applied and Environmental Biology,2010,16(4):577-580.(in Chinese)
[2]XU F,YANG Z F,CHEM B,etal.Impactof submerged plants on ecosystem health of the plant-dominated Baiyangdian Lake,China[J].EcologicalM odelling,2013,252(10):167-175.
[3]URUCPK,DEM IREZEN Y D.Response ofantioxidantdefences to Zn stress in three duckweed species[J].Ecotoxicology and Envtronmental Safety,2012,85:52-58.
[4]KOLADA A.Theeffectof lakemorphology on aquatic vegetation developmentand changesunder the influenceofeutrophication[J].Ecological Indicators,2014,38(0):282-293.
[5]李玥.镉、铜、锌对四种水生植物的毒性效应[D].长春:东北师范大学,2007.
[6]陈辉,施国新,徐勤松,等.Cr6+对菹草叶绿素荧光参数,抗氧化系统和超微结构的胁迫影响[J].植物研究,2009(5):559-564. CHEN Hui,SHIGuoxin,XU Qinsong,eta l.Toxic effectsof Cr6+on chlorophyll fluorescence parameters,antioxidantsystems and ultrastructureof potamogeton crispus[J].Bulletin of Botanical R esearch,2009,16(4):559-564.(in Chinese)
[7]贾荣.重金属汞对菹草无菌苗的毒害机理研究[D].南京:南京师范大学,2012.
[8]白宝章,汤学军.植物生理学测试技术[M].北京:中国科学技术出版社,1994.
[9]王学奎.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006.
[10]STOBARTA K,GRIFFITHSW T,AMEEN-BUKHARII,etal.Theeffectof Cd2+on thebiosynthesisof chlorophyll in leavesof barley[J].Physiologia Plantarum,1985,63(3):293-298.
[11]PAWLIK-SKOWRO?SKA B.Phytochelatin production in freshwater algae Stigeoclonium in response to heavymetals contained inm iningwater;effectsof some environmental factors[J].Aquatic Toxicology,2001,52(3-4):241-249.
[12]王君,徐勤松,韦淑丽,等.Zn2+在菹草叶片中的积累及其植物毒理学效应[J].西北植物学报,2009(11):2249-2255. WANG Jun,XU Qinsong,WEIShuli,etal.Zinc Accumulation and itseffectson physiologicaldynam icsand calcium distribution in submergedmacrophyte,Potamogeton crispus L.[J].Acta Bot Boreal-Occident Sin,2009(11):2249-2255.(in Chinese)
[13]徐勤松,施国新,许丙军,等.Cu、Zn在黑藻叶片中的富集及其毒理学分析[J].水生生物学报,2007(1):1-8. XU Qinsong,SHIGuoxin,XU Binjun,etal.Bioaccumulation and toxicity of Cu and Zn in Hydrilla verticillata(Linn.f.)royle[J]. Acta Hydrobiologica Sinica,2007(1):1-8.(in Chinese)
[14]徐勤松,施国新,计汪栋,等.Zn对荇菜叶片保护酶活性、渗透调节物质含量和Ca~(2+)定位分布的影响[J].水生生物学报,2009(4):613-619. XU Qinsong,SHI Guoxin,JIWangdong,et al.Effect of Zn on protective enzymes activity,osmolytes content and Ca2+distribution in leavesof Nymphoidespeltatum Zn[J].Acta Hydrobiologica Sinica,2009(4):613-619.(in Chinese)
[15]ALIM B,VAJPAYEE P,TRIPATHIR D,et al.Mercury bioaccumulation induces oxidative stress and toxicity to submerged macrophyte Potamogeton crispus L.[J].Bulletin of Environmental Contam ination and Toxicology,2000,65(5):573-582.
[16]李朝苏,刘鹏,徐根娣,等.铝浸种对荞麦种子萌发和幼苗生理的影响[J].生态学报,2006(6):2041-2047. LIChaosu,LIU Peng,XUGendi,etal.Effect of seed soaking w ith alum inum on seed germination and seedling physiology of buckwheat[J].Acta Ecologica Sinica,2006(6):2041-2047.(in Chinese)
[17]严政,谢水波,李仕友,等.凤眼莲、大薸对铀胁迫的生理生化响应[J].安全与环境学报,2012(3):1-5. YAN Zheng,XIEShuibo,LIShiyou,etal.Water hyacinth,Pistia stratiotes on the physiological response of uranium [J].Journal of Safety and Environment,2012(3):1-5.(in Chinese)
[18]湛灵芝,铁柏清,曾桂华.Cd2+、乙草胺单一及其复合污染对少根紫萍生理生化特性的影响[J].安全与环境学报,2013(3):9-13. ZHAN Linzhi,TIE Baiqing,ZENG Guihua.Individual and complex effects of Cd2+,ethyl am ine on the physiological characteristicsof spirodela oligorrhiza[J].Journal of Safety and Environment,2013(3):9-13.(in Chinese)
[19]姜岩.重金属铜对菹草无菌苗的毒理学研究[D].南京:南京师范大学,2013.
[20]杨红玉,王焕校.绿藻的镉结合蛋白及其耐镉性初探[J].植物生理学报,1985(4):357-365. YANG Hongyu,WANG Huanxiao.A prelim inary study on cadm ium-binding proteins of two green algae and their cadmium tolerance[J].Acta Phytophysiologica Sinica,1985(4):357-365.(in Chinese)
[21]ANWESHAM Bhaduri,M H Fulekar.Antioxidantenzyme responsesofplants to heavymetalstress[J].Reviews in Environmental Science and Biotechnology,2012,11(1):55-69.
[22]计汪栋,施国新,张慧,等.菹草对Hg2+胁迫的生理和结构应答反应[J].生态学报,2007(7):2856-2863. JIWangdong,SHIGuoxin,ZANG Hui,etal.Physiologicaland ultrastructural responses of Potamogeton crispus to Hg2+stress[J]. Acta Ecologica Sinica,2007(7):2856-2863.(in Chinese)
科技信息
日本发现含有贝壳钙的除菌剂可以防止食物中毒
2016年7月12日,据日媒报道,日本京大等研究组发表消息称,肉食加工中的生拌用牛肉使用融入贝壳钙的除菌剂可以防止食物中毒。但是,目前日本限制在店铺销售生拌牛肉用生肉,研究组致力于放宽规制,将其实用化。
据研究组称,将生肉块在除菌剂中浸5~10分钟、使用像洗衣机那样的机械高速洗净后,附着在生肉上的病原性大肠菌0157与处理前相比,至少减少至1%以下。
因销售生肉,不断出现食物中毒而死亡的事例,日本不断强化肉食相关的规制。目前,店铺上提供的生拌用牛肉,只能是提供经过一次加热杀菌处理而形成的“削减的肉块”。
京大研究组的教授称,如果使用除菌的方法,与加热处理相比,可以减少“肉的削减量”,今后可能提供更便宜的生拌牛肉用生肉。
[信息来源]食品伙伴网.日本发现含有贝壳钙的除菌剂可以防止食物中毒 [EB/OL].(2016-7-12).http://news. foodmate.net/2016/07/387259.html
Stress Effect and Response M echanism of Cd2+,Cu2+,Zn2+and Pb2+on Potamogeton crispus L.Grow th
LIWanlu1,2,ZHANG Guangsheng1,2,CHENG Xiaoying*1,2
(1.School of Environment andCivil Engineering,JiangnanUniversity,Wuxi 214122,China;2.Jiangsu Coorperative Innovation Center of Water Treatment Technology and Material,Suzhou 215000,China)
To estimate the influence of different heavy metals to the Potamogeton crispus L.,especially the grow th and development of itsw inter buds,the stresses of Cd2+,Cu2+,Zn2+and Pb2+in different concentrations to the grow th and development of w inter buds of Potamogeton crispus L. were studied,through the quantitative determ ination of germ inated number of w inter buds,chlorophyll of the leaves,soluble protein,malondialdehyde(MDA),and antioxidant system.As a result,under the same concentrations,Cu2+had the most obvious effect on physiological and biochemical indexes of the plant.The Cd2+ranked the second and followed by Pb2+and Zn2+.The plants cannotgrow normally oreven survivew ith concentration of Cu2+or Cd2+higher than 0.5mg/L. At the same time,the plants can grow normally under 10 mg/L of Zn2+or Pb2+treatment,which provided effectivebasis foraquatic plant in themetal threshold forecological repair.
stress,Potamogeton crispus L.,Cd2+,Cu2+,Zn2+,Pb2+
Q 948
A
1673—1689(2016)09—1001—07
2015-01-05
国家重大水专项项目(2012ZX07101-013-04)。
成小英(1977—),女,江苏无锡人,理学博士,副教授,硕士研究生导师,主要从事水环境治理及其机理方面的研究。E-mail:chengxiaoysytu@163.com
猜你喜欢
肝博士(2022年3期)2022-06-30
海外星云(2021年9期)2021-10-14
阅读(科学探秘)(2020年8期)2020-11-06
中国果业信息(2019年1期)2019-01-05
中成药(2018年8期)2018-08-29
生物学教学(2017年9期)2017-08-20
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年9期)2015-03-01
现代检验医学杂志(2015年6期)2015-02-06
食品工业科技(2014年6期)2014-05-10
