
稀土元素铈对毛葱生理活性和根尖细胞有丝分裂的影响
2016-09-07 00:54:14王亚喆徐秋曼天津师范大学生命科学学院天津市动植物抗性重点实验室天津300387
天津师范大学学报(自然科学版) 2016年1期
刘 慧,王亚喆,徐秋曼(天津师范大学 a.生命科学学院,b.天津市动植物抗性重点实验室,天津 300387)
稀土元素铈对毛葱生理活性和根尖细胞有丝分裂的影响
刘慧,王亚喆,徐秋曼
(天津师范大学 a.生命科学学院,b.天津市动植物抗性重点实验室,天津 300387)
摘要:为探究稀土元素铈对毛葱幼苗生理活性和根尖细胞有丝分裂的影响,采用不同质量浓度的Ce(NH4)2(NO3)6培养液水培毛葱幼苗,测量毛葱幼苗根和叶的生长、相对电导率、丙二醛和叶绿素含量,观察根尖细胞的染色体畸变情况.结果表明:1mg/L和10mg/L的Ce(NH4)2(NO3)6处理能够显著促进毛葱幼苗根和叶的生长,提高幼苗的叶绿素含量,降低幼苗的丙二醛含量和相对电导率,其中1mg/L的Ce(NH4)2(NO3)6作用最显著.50mg/L和100mg/L的Ce(NH4)2(NO3)6则抑制了毛葱根和叶的生长,使叶绿素含量降低,提高了丙二醛含量和相对电导率,其中100mg/L的Ce(NH4)2(NO3)6抑制作用最强烈.Ce(NH4)2(NO3)6质量浓度越大,毛葱根尖细胞的染色体畸变率越大,低质量浓度的Ce(NH4)2(NO3)6处理下,根尖细胞会出现可逆的染色体桥现象,高浓度处理下,根尖细胞会出现不可逆的染色体断片和粘连现象.
关键词:毛葱;Ce(NH4)2(NO3)6;生理活性;染色体畸变
适量的稀土元素对植物有益,表现为促进植物生长发育、提高植物光合作用能力和抗逆能力等[1-3].余海兵等[4]研究农用稀土对糯玉米幼苗光合与生理指标的影响时发现,施用农用稀土能够显著提高幼苗的光合参数、根系活力和抗氧化酶活性;周青等[5]研究发现,稀土元素镧能够缓解土壤重金属镉对玉米幼苗的毒害作用;赵朝宇等[6]也发现铈能够增强黄豆幼苗抗铅胁迫的能力.高浓度的稀土元素对植物也会产生毒害作用,主要表现为根和叶的生长速度减缓、叶片枯萎、变黄及其他生理生化指标弱化等.如赵倩倩等[7]的研究表明,采用质量浓度50 mg/L的硝酸铵铈溶液处理黄豆种子能够明显抑制种子萌发和根系活力,硝酸铵铈溶液浓度增大,对黄豆种子萌发和幼苗的抑制作用增强.稀土元素在植物不同部位的富集程度也有差异,根是富集稀土元素最多的部位;其次是叶、茎;作物的种子和果实中富集的稀土元素最少[8].
毛葱(Allium cepa var.agrogarum L.)为百合科(Liliaceae)葱属(Allium)洋葱(Allium cepa)的变种,营养价值高,同时有很好的医疗作用.毛葱的适应能力非常强,易于培养而且生长周期短,是非常好的实验材料[9].本文以毛葱为材料,采用营养液培养法,研究不同浓度的稀土元素铈处理下毛葱生长和生理指标的情况,以及根尖细胞有丝分裂过程中染色体形态发生的变化,以此探究铈对毛葱生理活性的影响.
1 材料与方法
1.1材料培养和处理
选取大小一致、未萌发的健康毛葱50个.在室温25℃条件下,剥去毛葱外面的干燥膜质,平均分成5组,利用水培方法进行培养,每24 h换1次水.3 d后改用Hoagland全营养液(每4 d更换1次)继续培养12 d.培养毛葱所用的Hoagland营养液分为4组,加入不同质量的Ce(NH4)2(NO3)6,使其最终质量浓度分别为1、10、50和100 mg/L,剩余1组作为对照组.在后续的12 d培养过程中对毛葱幼苗的生长指标、相对电导率、MDA含量和叶绿素含量进行测定,同时对毛葱根尖进行切片,观察其染色体形态的变化.
1.2测定方法
毛葱幼苗经不同质量浓度的Ce(NH4)2(NO3)6培养液处理后,每组随机选取6株幼苗,每隔24 h对根长和叶长进行测定.相对电导率测定:按照仲强等[10]的方法,20℃下测定幼苗电导,之后煮沸5min,冷却至20℃时再测电导,二者比值为相对电导率.MDA含量测定:按照成广雷等[11]的方法,单位用μmol/g表示.叶绿素含量测定:处理10 d后利用丙酮提取法测定叶绿素含量.根尖细胞染色体形态观察:在每一个处理组中随机取5个毛葱根尖,用Carnoy′s固定液固定,常规方法压片,卡宝品红染色[12].
1.3数据处理
实验数据采用Spss17.0处理,采用Excel 2003软件作图.
2 结果与分析
2.1铈对毛葱幼苗根和叶片生长的影响
不同质量浓度的Ce(NH4)2(NO3)6培养液处理下,毛葱幼苗根和叶片的生长情况如图1所示.
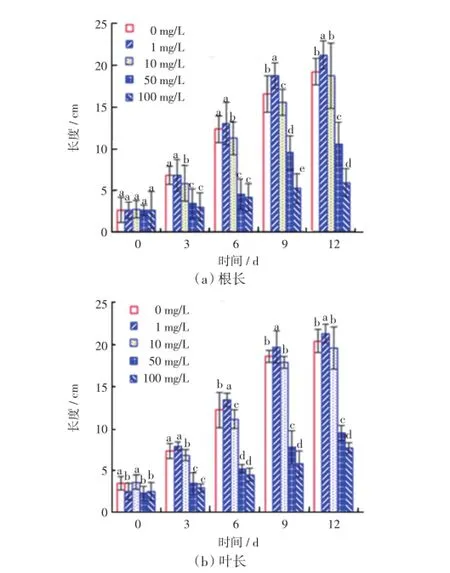
图1 不同质量浓度的Ce(NH4)2(NO3)6溶液培养下毛葱的根长和叶长Fig.1 Root length and leaf length of Allium cepa var. agrogarum L.cultured in Ce(NH4)2(NO3)6solutions with different mass concentrations
由图1(a)可以看出,随着培养时间的延长,5个处理组中毛葱的根都有生长,但不同处理组中长势差异明显.对照组毛葱的根生长速度较快、根系发达、根呈白色;1 mg/L的Ce(NH4)2(NO3)6处理组中毛葱根的长势好于对照组,尤其是在培养9 d以后,同对照组之间的差异具有统计学意义(p≤0.05);10 mg/L的Ce(NH4)2(NO3)6处理组中根的长势比对照组略有下降,培养时间越长,同对照组的差异越大,而且根呈现淡黄色;当Ce(NH4)2(NO3)6的质量浓度进一步增大至50 mg/L时,毛葱根的长势明显变差,2个高质量浓度处理组的根长显著低于对照组和低质量浓度处理组,差异具有统计学意义(p≤0.05),尤其是100 mg/L的处理组,根系生长受到较强抑制,毛葱根生长缓慢、稀疏,根呈黄色,后期出现腐烂现象.
由图1(b)可以看出,5个处理组中毛葱叶片长势差异与根的情况基本相同.1 mg/L的Ce(NH4)2(NO3)6处理组中,叶片长势最好,叶长在培养6 d后显著高于对照组和其他处理组;对照组叶片长度在培养前期高于10、50和100 mg/L的Ce(NH4)2(NO3)6处理组,6 d以后,对照组与10 mg/L的Ce(NH4)2(NO3)6处理组叶片长度相近;剩余2个处理组中,毛葱长势较差,叶片长度显著低于对照组和低质量浓度处理组的数值,差异具有统计学意义(p≤0.05).由此可见,1 mg/L 的Ce(NH4)2(NO3)6处理对毛葱叶片生长有促进作用,而高浓度的2个处理组中毛葱叶片生长则受到较强的抑制作用,且颜色呈现黄色.
2.2铈对毛葱幼苗相对电导率的影响
相对电导率能够反映植物膜系统的状况,植物在逆境或受到毒害的情况下,膜系统会受到损伤,生物膜透性增加,细胞膜容易破裂,使细胞内一些可溶性物质外渗,从而电导率增大[13-15].本研究中用不同质量浓度的Ce(NH4)2(NO3)6培养液培养毛葱,其相对电导率的变化如图2所示.
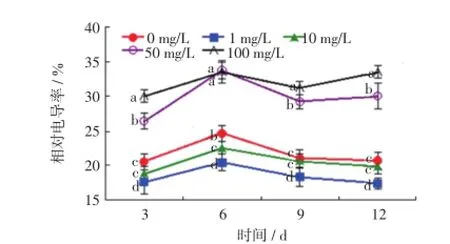
图2 不同质量浓度的Ce(NH4)2(NO3)6溶液培养下毛葱幼苗的相对电导率Fig.2 Relative conductivity of Allium cepa var.agrogarum L. cultured in Ce(NH4)2(NO3)6solutions with different mass concentrations
由图2可以看出,随着培养时间的延长,不同处理组中毛葱幼苗的相对电导率均呈现上升趋势,并且在培养第6天,各组的毛葱幼苗相对电导率均达到峰值.但在相同培养时间内,质量浓度≤10 mg/L的Ce(NH4)2(NO3)6能够降低毛葱幼苗的相对电导率,尤其是1mg/L的处理组中相对电导率显著低于对照组和10 mg/L处理组,差异具有统计学意义(p≤0.05).高浓度处理组中毛葱幼苗的相对电导率则显著高于对照组和低浓度处理组,尤其是100 mg/L的处理组,相对电导率超过了30%.
2.3铈对毛葱幼苗MDA含量的影响
植物在逆境条件或者器官衰老阶段,细胞膜脂质中的不饱和脂肪酸会发生过氧化作用,MDA大量产生,最终导致膜透性增大.因此MDA含量可以作为衡量细胞膜脂质过氧化程度的指标[16].用不同质量浓度的Ce(NH4)2(NO3)6培养液培养毛葱,根和叶片中的MDA含量如图3所示.
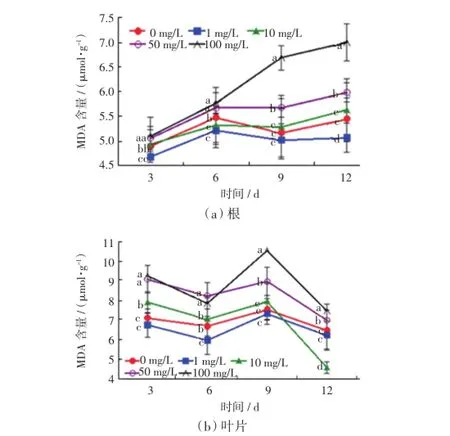
图3 不同质量浓度的Ce(NH4)2(NO3)6溶液培养下毛葱幼苗的MDA含量Fig.3 MDA content of Allium cepa var.agrogarumL. cultured in Ce(NH4)2(NO3)6solutions with different mass concentrations
由图3可以看出,在12 d的培养过程中,只有1 mg/L的Ce(NH4)2(NO3)6处理组中毛葱根和叶片的MDA含量始终低于对照组.10 mg/L的处理组中MDA含量与对照组接近;Ce(NH4)2(NO3)6的质量浓度高于50 mg/L时,毛葱根和叶片的MDA含量都显著高于对照组,差异具有统计学意义(p≤0.05),质量浓度为100 mg/L的处理组中,MDA的含量最大.由此可见,低浓度的铈可以降低细胞中MDA的含量,即降低质膜通透性;高浓度的铈则会增加细胞中MDA的含量,增加质膜通透性.
2.4铈对毛葱幼苗叶绿素含量的影响
测定不同质量浓度的Ce(NH4)2(NO3)6处理下毛葱幼苗叶绿素的含量,结果如图4所示.由图4可以看出,随着培养时间的延长,对照组和低质量浓度的Ce(NH4)2(NO3)6溶液处理组中,毛葱幼苗的叶绿素含量均呈现增加趋势;50 mg/L和100 mg/L的处理组中,幼苗的叶绿素含量则随着培养时间的延长呈现下降趋势.相同时间下,质量浓度为1mg/L的处理组中毛葱幼苗的叶绿素含量始终显著高于其他4个处理组,差异具有统计学意义(p≤0.05);其次是质量浓度为10 mg/L的处理组,同对照组相近;50mg/L和100mg/L的处理组中毛葱幼苗的叶绿素含量显著低于对照组,其中100 mg/L的处理组中毛葱幼苗的叶绿素含量最低.由此可见,低浓度的铈能够促进毛葱的叶绿素合成,高浓度的铈则有抑制作用.

图4 不同质量浓度的Ce(NH4)2(NO3)6溶液培养下毛葱幼苗的叶绿素含量Fig.4 Chlorophyll content of Allium cepa var.Agrogarum L. cultured in Ce(NH4)2(NO3)6solutions with different mass concentrations
2.5铈对毛葱根尖细胞有丝分裂的影响
2.5.1铈对毛葱根尖细胞染色体畸变率的影响
采用不同质量浓度的Ce(NH4)2(NO3)6溶液处理毛葱,其根尖细胞染色体产生畸变的情况如表1所示.
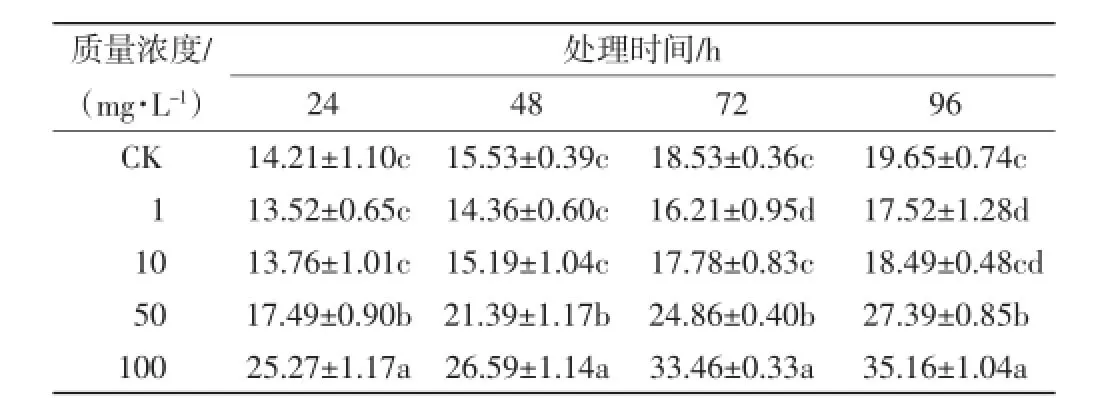
表1 不同质量浓度的Ce(NH4)2(NO3)6溶液培养下毛葱根尖细胞染色体的畸变率Tab.1 Chromosomal aberrations of root tip cells of Allium cepa var.agrogarum L.cultured in Ce(NH4)2(NO3)6solutions with different mass concentrations
从表1可以看出,随着培养时间的延长,5个处理组中毛葱根尖细胞染色体的畸变率均增加.质量浓度为l mg/L的Ce(NH4)2(NO3)6溶液处理组中,根尖细胞染色体的畸变率低于对照组和其他处理组,尤其是培养72 h时,同其他4组之间的差异具有统计学意义(p≤0.05);质量浓度为l0mg/L的Ce(NH4)2(NO3)6溶液处理组中,染色体畸变率略低于对照组,显著低于高浓度处理组,差异具有统计学意义(p≤0.05);随着Ce(NH4)2(NO3)6溶液质量浓度的增大,毛葱根尖细胞染色体的畸变率上升,质量浓度为100 mg/L的处理组中畸变率显著高于同时段其他处理组的畸变率,差异具有统计学意义(p≤0.05).
2.5.2铈对毛葱根尖细胞染色体形态的影响
观察毛葱根尖细胞的有丝分裂,结果如图5所示.

图5 毛葱根尖细胞的有丝分裂Fig.5 Mitosis of root tip cells of Allium cepa var. agrogarum L.
在经过不同质量浓度的Ce(NH4)2(NO3)6处理后,毛葱根尖细胞的染色体可能出现畸变,如染色体断片(chromosomalfragment)、染色体桥(chromosomebridge)、染色体粘连(chromosome sticky)等.染色体断片指的是一个染色体发生一处或多处断裂而不重接且远远分开所出现的一个或多个无着丝粒的节段,主要发生在细胞分裂中期.染色体桥出现在有丝分裂后期,当2个染色单体的着丝粒己经分别移向相对的两极后,二者的臂仍然粘连在一起,形成后期桥.染色体粘连表现为染色体相互粘连并且聚缩成不规则的团块状[17].采用10mg/L的Ce(NH4)2(NO3)6溶液处理毛葱96h后,在细胞有丝分裂后期会出现染色体双桥现象,如图5(c)和5(d)所示,用50 mg/L的Ce(NH4)2(NO3)6溶液处理毛葱48 h也会出现这一现象,而且Ce(NH4)2(NO3)6浓度越高染色体畸变越明显,这是一种可逆的中度毒害作用,恢复正常后其染色体结构及数目会发生变化. 用100 mg/L的Ce(NH4)2(NO3)6处理毛葱24 h后毛葱根尖细胞出现染色体断片现象,48 h后会出现染色体粘连现象,如图5(e)和5(f)所示,这2种都是不可逆的能够导致细胞致死的毒害作用.
3 讨论与结论
本研究采用不同质量浓度的Ce(NH4)2(NO3)6溶液对毛葱幼苗进行培养,测量幼苗的生长及生理指标,并且观察毛葱根尖细胞的有丝分裂情况.结果发现,铈对于毛葱的生长具有低促高抑现象.质量浓度为1mg/L的处理组中,根和叶片的生长、叶片叶绿素含量都好于对照组和高浓度处理组,相对电导率和MDA含量则低于其他组;Ce(NH4)2(NO3)6的质量浓度高于50 mg/L时,毛葱幼苗的生长受到明显抑制.观察毛葱根尖细胞的有丝分裂,发现染色体的畸变率随着Ce(NH4)2(NO3)6处理浓度的增加而增大,而且低质量浓度的处理下细胞有丝分裂会出现可逆的染色体桥现象,高浓度处理下有丝分裂则出现不可逆的染色体断片和染色体粘连现象,这可能是高浓度铈抑制根部生长的根本原因.
参考文献:
[1]BRIOSCHI L,STEINMANN M,LUCOT E,et al.Transfer of rare earth elements(REE)from natural soil to plant systems:implications for the environmental availability of anthropogenic REE[J].Plant Soil,2013,366(1/2):143-163.
[2]金姝兰,黄益宗.稀土元素对农田生态系统的影响研究进展[J].生态学报,2013,33(16):4836-4845.
JIN S L,HUANG Y Z.A review on rare earth elements in farmland ecosystems[J].Acta Ecologica Sinica,2013,33(16):4836-4845(in Chinese).
[3]CARPENTER D,BOUTIN C,JANE E,et al.Uptake and effects of six rare earth elements(REEs)on selected native and crop species growing in contaminated soils[J].PLoS ONE,2015,10(6):e0129936.
[4]余海兵,刘正,王波.农用稀土对糯玉米幼苗光合变化和生理指标的分析[J].中国稀土学报,2011,29(1):119-124.
YU H B,LIU Z,WANG B.Analysis of rare earths in agriculture on photosynthetic changes and physiological indicators in waxy corn seedlings[J].Journal of the Chinese Rare Earth Society,2011,29(1):119-124(in Chinese).
[5]周青,陆敢超,张辉,等.稀土La对Cd胁迫下玉米幼苗生长影响[J].农业环境科学学报,2004,23(1):18-21.
ZHOU Q,LU G C,ZHANG H,et al.Effects of La on the growth of corn seedling under Cd stress[J].Journal of Agro-Environment Science,2004,23(1):18-21(in Chinese).
[6]赵朝宇,刘慧,王亚喆,等.稀土元素铈对黄豆幼苗铅胁迫的缓解效应[J].天津师范大学学报:自然科学版,2015,35(4):76-80.
ZHAO C Y,LIU H,WANG Y Z,et al.Mitigative effect of cerium on Pb stress in soybean seedlings[J].Journal of Tianjin Normal University:Natural Science Edition,2015,35(4):76-80(in Chinese).
[7]赵倩倩,赵朝宇,徐秋曼,等.稀土金属铈对黄豆种子萌发及幼苗生理活性的影响[J].天津师范大学学报:自然科学版,2014,34 (4):72-75. ZHAO Q Q,ZHAO C Y,XU Q M,et al.Effects of cerium on germination of soybean seeds and physiological activity of seedlings[J].Journal of Tianjin Normal University:Natural Science Edition,2014,34(4):72-75(in Chinese).
[8]XU X K,ZHU W Z,WANG Z J,et al.Distribution of rare earths and heavy metals in field-grown maize after application of rare earth-containing fertilizer[J].Sci Total Environ,2002,293(1/2/3):97-105.
[9]邹金华,张忠贵,魏爱琪.毛葱的镉吸收积累及亚细胞分布特征[J].天津师范大学学报:自然科学版,2014,34(4):74-79. ZHOU J H,ZHANG Z G,WEI A Q.Characteristics of accumulation and subcellular distribution of cadium in Allium cepa var.agrogarum[J]. Journal of Tianjin Normal University:Natural Science Edition,2014,34(4):74-79(in Chinese).
[10]仲强,康蒙,郭明,等.浙江天童常绿木本植物的叶片相对电导率及抗寒性[J].华东师范大学学报:自然科学版,2014,34(4):72-75. ZHONG Q,KANG M,GUO M,et al.Leaf frost sensitivity and percentage of electrolyte leakage of the evergreen woody species in Tiantong region,Zhejiang Province[J].Journal of East China Normal University:Natural Science,2014,34(4):72-75(in Chinese).
[11]成广雷,张海,赵久然,等.临界胁迫贮藏条件下不同基因型玉米种子活力及生理变化[J].中国农业科学,2015,48(1):33-42. CHENG G L,ZHANG H,ZHAO J R,et al.Vigor and physiological changes of different genotypes of maize seed(Zea mays L.)under critical stress storage conditions[J].Scientia Agricultura Sinica,2015,48 (1):33-42(in Chinese).
[12]王晓涧,李娜,陈宗礼.洋葱根尖染色体标本的制备方法[J].延安大学学报:自然科学版,2007,26(2):73-74. WANG X J,LI N,CHEN Z L.A new method for chromosome preparation of Aillum cepa root point[J].Journal of Yan′an University:Natural Science Edition,2007,26(2):73-74(in Chinese).
[13]张燕,方力,李天飞,等.钙对低温胁迫的烟草幼苗某些酶活性的影响[J].植物学通报,2002,19(3):342-347. ZHANG Y,FANG L,LI T F,et al.Effect of Ca2+on activities of some enzymes in tobacco seedlings under cold stress[J].Chinese Bulletin of Botany,2002,19(3):342-347(in Chinese).
[14]徐秋曼,元英进,程景胜,等.稀土元素铈对红豆杉细胞膜透性的影响[J].稀土,2004,25(2):50-53. XU Q M,YUAN Y J,CHENG J S,et al.Cell membrane permeability changes induced by Ce in suspension cultures of taxus cuspidate[J]. Chinese Rare Earths,2004,25(2):50-53(in Chinese).
[15]李庆,王应军,宗贵仪,等.铈缓解镉对玉米种子的毒害效应研究[J].稀土,2013,34(6):1-6. LI Q,WANG Y J,ZONG G Y,et al.Moderating effect of cerium on toxic action of cadium to corn seed[J].Chinese Rare Earths,2013,34 (6):1-6(in Chinese).
[16]XU Q M,CHEN H.Antioxidant responses of rice seedling to Ce4+under hydroponic cultures[J].Ecotoxicology and Environmental Safety,2011,74(6):1693-1699.
[17]周锦雯.镉对蚕豆根尖细胞有丝分裂的影响[D].成都:四川师范大学,2006. ZHOU J W.Effects of Cd2+on cell division of root tips of Vicia faba[D]. Chengdu:Sichuan Normal University,2006(in Chinese).
(责任编校纪翠荣)
第一作者:刘慧(1991—),女,硕士研究生.
文章编号:1671-1114(2016)01-0052-05
中图分类号:Q945
文献标志码:A
收稿日期:2015-09-12
基金项目:天津市自然科学基金资助项目(13JCYBJC25500).
通信作者:徐秋曼(1969—),女,副教授,主要从事植物生理学方面的研究.
Effects of cerium on physiological activity of seedlings and mitosis of root tip cells in Allium cepa var.agrogarum L.
LIU Hui,WANG Yazhe,XU Qiuman
(a.College of Life Sciences,b.Tianjin Key Laboratory of Animal and Plant Resistance,Tianjin Normal University,Tianjin 300387,China)
Abstract:To study the effect of cerium on physiological activity of seedlings and mitosis of root tip cells in Allium cepa var.agrogarum L.,the seedlings were hydroponic cultivated with different mass concentrations of Ce(NH4)2(NO3)6solutions. Several physiological characteristics were measured,including root and leaf lengh,relative conductivity,MDA and chlorophyll content as well as chromosomal aberration of root tip cells.The results showed that the Ce(NH4)2(NO3)6solutions with mass concentrations of 1 mg/L and 10 mg/L can promote the growth of roots and leaves,increase the chlorophyll content,and reduce the MDA content and relative conductivity.And 1 mg/L of Ce(NH4)2(NO3)6solution had the most significant promoting effect.As for the Ce(NH4)2(NO3)6solutions with high mass concentrations(50 mg/L and 100 mg/L),they can inhibit the growth of the roots and leaves,decrease the chlorophyll content,and increase the MDA content and relative conductivity. The treatment of 100 mg/L of Ce(NH4)2(NO3)6had the most significant inhibiting effect.With the increase of mass concentrations of Ce(NH4)2(NO3)6solutions,the frequencies of chromosomal aberration of root tip cells increased as well.The low mass concentrations of Ce(NH4)2(NO3)6solutions can induce the reversible chromosome bridge,while the high mass concentrations can result in the irreversible chromosomal fragment and chromosome sticky.
Keywords:Allium cepa var.agrogarum L.;Ce(NH4)2(NO3)6;physiological activity;chromosomal aberration
猜你喜欢
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
科学之谜(2019年3期)2019-03-28 10:29:44
中国果业信息(2019年1期)2019-01-05 17:41:42
科学之谜(2018年8期)2018-09-29 11:06:46
浙江工业大学学报(2017年5期)2018-01-22 02:03:40
现代园艺(2017年23期)2018-01-18 06:57:46
生物学教学(2017年9期)2017-08-20 13:22:32
恋爱婚姻家庭·养生版(2016年9期)2016-09-07 11:25:01
中央民族大学学报(自然科学版)(2015年2期)2015-06-09 08:45:16
食品工业科技(2014年6期)2014-05-10 06:04:50
