
香草兰VpLFY基因的克隆与表达分析
2016-05-30 10:48:04顾文亮王辉庄辉发王华朱自慧宋应辉
热带农业科学 2016年2期
顾文亮 王辉 庄辉发 王华 朱自慧 宋应辉

摘 要 花分生组织特征基因LEAFY在植物花芽分化与成花诱导途径中起着重要的调控作用。通过RACE方法从香草兰中克隆VpLFY基因的全长cDNA序列。结果显示,该基因包含一个1 493 bp的开放阅读框,编码491个氨基酸残基。经蛋白质序列同源比对分析结果发现,VpLFY属于FLO-LFY家族。组织特异性研究结果表明,VpLFY基因在香草兰组织中属于组成型表达。荧光定量分析结果表明,VpLFY基因分别在香草兰纯花芽和混合花芽的不同发育阶段中有着较强的表达。
关键词 香草兰 ;VpLFY ;基因克隆 ;表达分析
分类号 S682.31 Doi:10.12008/j.issn.1009-2196.2016.02.006
Molecular Cloning and Expression of VpLFY Gene from Vanilla
(Vanilla planifolia Andrews)
GU Wenliang1,2) WANG Hui1,3) ZHUANG Huifa1,3)
WANG Hua1,3) ZHU Zihui1,3) SONG Yinghui1,2,3)
(1 Spice and Beverage Research Institute, CATAS, Wanning, Hainan 571533;
2 Key Laboratory of Genetic Resources Utilization of Spice and Beverage Crops,
Ministry of Agriculture, Wanning, Hainan 571533;
3 Hainan Provincial Key Laboratory of Genetic Improvement and Quality Regulation
for Tropical Spice and Beverage Crops, Wanning, Hainan 571533)
Abstract Floral meristem identity gene LEAFY is a master regulator of flower bud differentiation and floral induction. In this study, we report the molecular characteristics of LFY gene cloned from Vanilla(Vanilla planifolia Andrews) using a RACE-PCR based strategy. VpLFY contained an open reading frame of 1 493 bp which encoded a polypeptide of 491 amino acids. Protein alignment showed that VpLFY contained typical domains which belonged to FLO-LFY protein family. Tissue-specific studies showed that the expression of VpLFY was constitutive expressed in various tissues of Vanilla. Fluorescent quantitative PCR analysises indicated that the expression of VpLFY were increased in reproductive bud and mixed bud at different stages of Vanilla flower bud differentiation. These results suggest that VpLFY may play an important role in the regulation of Vanilla flower bud differentiation and floral induction.
Keywords Vanilla planifolia Andrews ; VpLFY ; gene clone ; expression analysis
花分生组织特征基因LEAFY(以下简称LFY基因)在成花相关基因的调控网络中处于关键位置[1],是植物从营养生长到生殖生长转变调节的中心,并且参与到多条花芽分化与成花诱导途径中,其表达始终贯穿于花序分化和成花发育的各个阶段,被认为是成花转变的标志性基因[2]。LFY基因不仅促进花分生组织的形成及其属性的调控,而且参与维持花分生组织的功能、成花基因的启动和防止花分生组织的逆转,同时参与花分生组织属性的决定及其进一步发育[3]。
香草兰(Vanilla planifolia Andrews)是兰科(Orchidaceae)香草兰属(Vanilla)热带攀援藤本植物,被誉为食品香料之王,广泛用于食品、饮料、化妆品和医药等行业中[4]。香草兰的花芽分化与成花诱导过程中存在着营养生长向生殖生长转变不彻底的现象,这极大影响着生产种植上香草兰的产量[5]。LFY基因在植物花芽分化与成花诱导途径中发挥着重要的作用。目前尚无香草兰VpLFY基因的相关研究报道,克隆香草兰VpLFY基因的全长序列并分析其功能,将有助于解析香草兰花芽分化与成花诱导的分子机理。本研究基于香草兰转录组测序信息,通过3′RACE和5′RACE方法从香草兰中克隆了一个VpLFY基因,采用荧光定量分析该基因的表达模式,探讨VpLFY基因在香草兰中的表达特征,有助于进一步解析香草兰花芽分化与成花诱导的调控机理。
1 材料与方法
1.1 材料
供试植株为生长3 a以上的墨西哥香草兰,种植在海南省万宁市南桥高龙的香料饮料研究所基地中,所有供试的香草兰植株均为荫蔽条件下露地栽培。总RNA的提取选取花芽分化初期的香草兰花芽组织。选择长势良好且大小一致的香草兰植株,选取花芽分化不同阶段的纯花芽、混合花芽和叶芽等芽体组织。对香草兰花芽分化进行连续观测至花分生组织分化,通过芽体组织的生长量及观测数据将香草兰花芽分化过程划分为6个时期,分别为花芽特征分化期(I)、花芽分化初期(II)、花芽分化中期(III)、花序分化初期(IV)、花序分化中期(V)、花分化初期(VI)[6]。在香草兰花芽分化的6个时期中,分别采集花芽、混合花芽和叶芽的芽体组织各6个待测样品,取样的芽体组织用液氮快速冷冻后放入-80℃超低温冰箱中贮存。取样的芽体组织用液氮快速冷冻后放入-80℃超低温冰箱中贮存。
1.2 方法
1.2.1 香草兰花芽组织总RNA的提取
香草兰花芽组织总RNA的提取采用RNeasy Plant Mini Kit(Qiagen)进行,具体提取步骤参照该试剂盒说明书,对提取后的总RNA进行分光光度计和电泳分析。
1.2.2 香草兰VpLFY基因的克隆与分析
按照香草兰的转录组测序结果设计3′RACE与5′RACE引物,3′RACE扩增的引物序列为LFY-F1(5′-CAAGTGCCCCACCAAGGTGACGAACC-3′)和LFY-F2(5′-CAAGCCCAAGATGCGGCATTACGTG-3′),5′RACE扩增的引物序列为LFY-R1(5′-GGTCCTTGGCGACGGCTTGGACTT-3′)和LFY-R2(5′-GAGGAGGAACTCGTGGCATTGCTCGTAG-3′)。按照GeneRacer Kit(Invitrogen)说明书进行RACE扩增反应。分别将3′RACE和5′RACE扩增得到的特异条带切胶回收,连接pLB Simple载体(天根),转化感受态细胞后送往生工生物工程(上海)股份有限公司进行测序。DNA序列经测序后在NCBI数据库BLASTn和蛋白数据库BLASTx中进行同源核苷酸和蛋白质序列检索。
1.2.3 香草兰VpLFY基因的表达分析
选择不同花芽分化阶段的叶芽、花芽与混合花芽等芽体组织,按照RNeasy Plant Mini Kit(Qiagen)的说明书分别提取总RNA,用RevertAidTM First Strand cDNA Synthesis Kit(Fermentas)合成cDNA第一链。以VpActin基因为内参基因,采用荧光定量PCR方法对VpLFY基因的组织特异性表达和不同分化阶段的表达特征进行分析。检测VpLFY基因的引物序列为VpLFY-F(5′-TGAGGGAGGAGGAGGTSGACGAYATGAT-3′)和VpLFY-R(5′-AGATBGAGAGGCGSGGATGSGCGTTGAA-3′)。内参基因VpActin的引物序列为VpActin-F(5′-GGGTTACTCCTTTACGACCACA-3′)和VpActin-R(5′-GGGTTACTCCTTTACGACCACA-3′)。采用SYBR Green荧光染料,荧光定量PCR的反应程序为:95℃预变性3 min,95℃变性7 s,60℃退火15 s,72℃延伸20 s,共40个循环。
1.2.3 数据处理与统计分析
每个实验处理均做3次生物学重复,采用Excel 2010软件处理实验数据与图表。
2 结果与分析
2.1 总RNA的提取
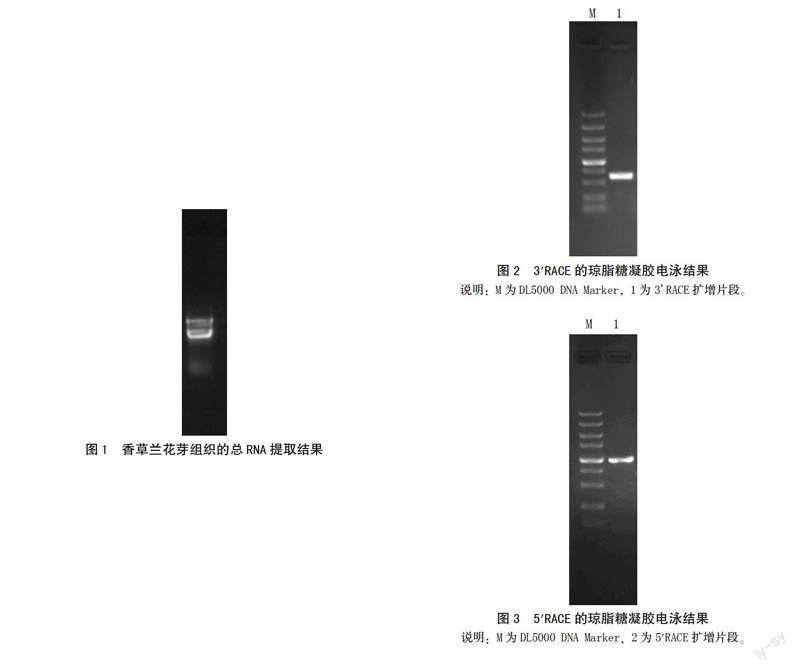
以1%的琼脂糖凝胶电泳检测提取的香草兰花芽组织总RNA(图1),28 S和18 S RNA的条带完整且边缘清晰,说明总RNA样品具有较好的完整性。经分光光度计检测,提取总RNA的OD260/OD280均在1.7~2.0,说明总RNA样品纯度较高。
2.2 VpLFY基因的克隆与序列分析
在香草兰花芽分化组织的转录组测序基础上,通过3′RACE扩增出一条特异性条带,大小约700 bp(图2),通过5′RACE扩增出一条特异性条带,大小约1 100 bp(图3)。将上述特异DNA片段经测序后拼接,通过BLAST比对发现该序列与其他植物LFY基因序列具有较高的同源性,表明该序列是香草兰LFY基因编码框全长cDNA,命名为VpLFY。VpLFY序列全长为1 708 bp,其中5′非编码区为38 bp,3′非编码区为197 bp,经分析可知VpLFY基因ORF长度为1 473 bp,编码491个氨基酸残基。根据生物信息学分析,预期分子量为54.13 ku,等电点为7.13。
在NCBI中对VpLFY蛋白质进行同源性分析,由图4可知,VpLFY蛋白与拟南芥、番茄、玉米和蝴蝶兰的LFY均存在同源性较高的保守氨基酸序列位点。另外,在NCBI中对VpLFY蛋白质保守结构域搜索,结果表明,VpLFY蛋白质属于FLO-LFY家族。
2.3 VpLFY基因的表达分析
采用荧光定量RT-PCR方法,以VpActin作为内参基因,分别对香草兰花芽分化不同组织中VpLFY基因的表达水平进行检测。由图5可知,VpLFY基因在所有香草兰花芽分化的组织中均有表达,其在纯花芽中表达量最高,在茎段和气生根中表达量较低。
采用荧光定量RT-PCR方法,以VpActin作为内参基因,分别对香草兰纯花芽、叶芽和混合花芽等芽体组织在不同分化阶段中VpLFY基因的表达水平进行检测。由图6可知,VpLFY基因在香草兰花芽分化的不同阶段中均有表达,其中纯花芽的VpLFY基因在花序分化初期(IV)表达量达到最高,混合花芽的VpLFY基因在花分化初期(VI)表达量达到最高,叶芽的VpLFY基因在整个花芽分化不同阶段表达量变化不大。
3 讨论与结论
LFY基因的首要功能是协同其他成花相关基因抑制分生组织细胞向营养生长发育[7]。研究结果发现,在成花转变前LFY基因主要表达于叶原基,当其表达量达一定阈值时,LFY基因则起抑制叶原基启动的作用,从而有助于分生组织细胞形成花原基[8]。在植物成花诱导中,LFY基因还参与维持花分生组织的正常功能、花启动、防止花分生组织的逆转[9]。在模式植物拟南芥中,LFY基因的表达最早出现于花序分生组织下侧花原基萌发的部位,但在花序分生组织中无表达。随着花分生组织的形成,LFY基因的表达量也逐渐增加,并分布于整个花分生组织中;当花器官原基开始分化出现时,LFY基因的表达大部分消失[10]。
本研究克隆了一个香草兰花分生组织特征基因VpLFY,该基因全长1 708 bp,包含一个1 473 bp的完整读码框,编码491个氨基酸残基。VpLFY编码的氨基酸序列的分子量为54.13 ku,等电点为7.13,该序列与其他植物LFY均存在同源性较高的保守位点,均属于FLO-LFY蛋白家族,这表明本研究克隆的VpLFY属于植物LFY基因。对VpLFY基因在香草兰中的组织特异性分析结果表明,香草兰花芽分化的纯花芽、混合花芽、叶芽、茎段、功能叶和气生根中均能检测到VpLFY基因的表达,这表明该基因在香草兰组织中属于组成型表达。对VpLFY基因在香草兰不同花芽分化阶段中的表达特征分析结果表明,香草兰的纯花芽在花芽分化前期均有较高的VpLFY基因表达,香草兰的混合花芽则在花芽分化后期有较高的VpLFY基因表达。根据上述结果推测,VpLFY基因可能在香草兰的花芽分化和成花诱导过程中发挥着重要的作用。
香草兰是兰科植物中最具经济价值的热带香料作物。已有研究结果表明,香草兰营养生长向生殖生长转变不彻底的现象极大影响着生产种植上香草兰的产量[11]。通过生物技术的手段为开展生产中的香草兰促花栽培措施是一直以来的目标[12]。本研究克隆的VpLFY基因及对其表达特征分析结果表明,香草兰花分生组织特征基因VpLFY可作为香草兰促花栽培措施的参考基因,通过检测VpLFY基因的表达强度可为生产上的生物技术促花栽培措施提供参考借鉴,这也是本研究下一步深入研究的方向。
参考文献
[1] Prenner G, Cacho N I, Baum D, et al. Is LEAFY a useful marker gene for the flower-inflorescence boundary in the Euphorbia cyathium?[J]. Journal of Experimental Botany, 2011, 62(1): 345-350.
[2] Weigel D, Alvarez J, Smyth D R, et al. LEAFY controls floral meristem identity in Arabidopsis[J]. Cell, 1992, 69(5): 843-859.
[3] Blazquez M A,Soowal L N,Lee I,et al. LEAFY expression and flower initiation in Arabidopsis[J]. Development, 1997, 124(19): 3 835-3 844.
[4] 王庆煌,朱自慧. 香草兰[M]. 北京:中国农业出版社,2004.
[5] 王 辉,庄辉发,王 华,等. 香草兰单株产量构成性状的通径分析[J]. 安徽农业科学,2010,38(29):16 198-16 199.
[6] 赵秋芳,陈娅萍,顾文亮,等. 香草兰花芽分化期蛋白质及碳水化合物变化研究[J]. 热带作物学报,2015,36(6):1 053-1 058.
[7] Jack T. Molecular and genetic mechanisms of floral control[J]. The Plant Cell, 2004, 16(Suppl):S1-S17.
[8] Moyroud E, Minguet E G, Ott F, et al. Prediction of regulatory interactions from genome sequences using a biophysical model for the Arabidopsis LEAFY transcription factor[J]. The Plant Cell, 2011, 23(4): 1 293-1 306.
[9] Winter C M, Austin R S, Blanvillain-Baufumé S, et al. LEAFY target genes reveal floral regulatory logic, cis Motifs, and a link to biotic stimulus response[J]. Development Cell, 2011, 20(4): 430-443.
[10] Blázquez M A, Ahn J H, Weigel D. A thermosensory pathway controlling flowering time in Arabidopsis thaliana[J]. Nature Genetics, 2003, 33(1):168-171.
[11] Havkin-Frenkel D, Belanger F C. Handbook of vanilla science and technology[M]. Southern Gate:Wiley-Blackwell, 2011.
[12] Anuradha K, Shyamala B N, Naidu M M. Vanilla-Its Science of cultivation, curing,chemistry, and nutraceutical properties[J]. Food Science and Nutrition, 2013, 53(12): 1 250-1 276.
猜你喜欢
中国中药杂志(2017年1期)2017-03-06 21:25:43
山东农业科学(2016年11期)2016-12-17 21:00:42
中国中药杂志(2016年20期)2016-11-19 12:27:25
江苏农业科学(2015年9期)2015-10-20 21:09:07
江苏农业科学(2015年5期)2015-10-20 21:08:26
江苏农业科学(2015年9期)2015-10-20 21:04:29
江苏农业科学(2015年8期)2015-09-10 20:57:43
江苏农业科学(2015年7期)2015-08-20 20:58:46
江苏农业科学(2015年4期)2015-06-15 21:04:08
南方农业·下旬(2015年1期)2015-04-16 01:10:20
