
胶孢炭疽菌漆酶基因Lac2的序列特征与表达分析
2015-10-20 21:09韦运谢等
江苏农业科学 2015年9期
韦运谢等

摘要:为了探明分离自芒果的胶孢炭疽菌(Colletotrichum gloeosporioides)漆酶同工酶基因的序列特征,进一步研究该菌的分子致病机理。以该菌为材料,采用同源克隆和RT-PCR法获得胶孢炭疽菌漆酶基因Lac2(KF924625)。结果表明,其编码区大小为1 785 bp,编码594个氨基酸,分子量为65.53 ku,等电点是6.56,含4个铜离子结合保守结构域;聚类分析发现,其预测蛋白与西瓜炭疽病菌(Colletotrichum orbiculare)的漆酶laccase-1(ENH77191.1)同源性最高,达到79%;半定量RT-PCR分析表明,在分生孢子不同萌发时段,Lac2的表达量有明显的升降趋势,接种后6 h表达量最低,9 h最高。可见Lac2具备真菌漆酶基因家族的序列特征,初步推测它可能与C. gloeosporioides分生孢子萌发侵染寄主有关,也可能参与调控芒果胶孢炭疽菌的漆酶活性和抗氧化反应等。
关键词:胶孢炭疽菌;Lac2基因;克隆;表达分析
中图分类号: Q785文献标志码: A文章编号:1002-1302(2015)09-0035-04
漆酶是含铜的多酚氧化酶,能够催化多种酚类化合物及其衍生物,使之生成相应的苯醌和水,且其催化底物具有广谱性,在环保、食品、医药、纺织等各个领域具有潜在的应用价值[1]。最早在漆树上发现漆酶,随后发现其也广泛存在于昆虫、高等植物、细菌及真菌中[2]。真菌漆酶与致病力、营养生长、增殖生长、孢子和黑色素的形成、氧化胁迫及渗透调节能力相关[3-5]。真菌中存在多种漆酶同工酶,且在不同生长期表达,功能也存在差异[6],如Colletotrichum orbiculare的Lac1与Lac2的功能截然不同,与野生型相比,Lac2突变体失去了致病力、黑色素含量下降及分生孢子颜色变浅,而Lac1突变体的黑色素含量略微减少、漆酶酶活却有所上升[7-8]。序列分析表明,不同漆酶基因氨基酸序列相似性不高,但在铜原子结合区域的保守性相当高,因此可以根据这些保守区设计简并引物同源克隆漆酶基因。
芒果(Mangifera indica L.)隶属漆树科芒果属,是世界五大著名热带水果之一。由胶孢炭疽菌(C. gloeosporioides)引起的芒果炭疽病是芒果最严重的病害之一,在整个生育期均可发病,且在贮运期危害最重,是限制芒果产业发展的主要因素之一。炭疽病菌的致病过程主要是通过病原菌分生孢子附着在寄主表面,萌发形成附着胞,附着胞内沉积一层黑色素,前端产生侵染钉侵入寄主并在寄主体内扩展,黑色素在病原菌穿透寄主组织中起着非常重要的作用,细胞壁降解酶起着软化细胞的辅助作用[9-10],而漆酶又是形成DHN黑色素必不可少的氧化酶[11],因此推测在芒果炭疽菌中漆酶可能通过干扰附着胞黑色素的合成而影响着该菌的致病力。目前,关于芒果胶胞炭疽菌的漆酶基因仅见笔者所在实验室报道的Lac1[12],为了系统研究各漆酶在芒果炭疽菌中的功能,本研究采用同源克隆策略,从芒果胶胞炭疽菌中又克隆获得了漆酶新基因Lac2,这为进一步研究其功能打下了材料基础。
1材料与方法
1.1供试菌株
芒果胶胞炭疽菌(Colletotrichum gloeosporioides)单孢菌株A2由中国热带农业科学院环境与植物保护研究所鉴定并提供。
1.2方法
1.2.1病原菌菌丝的收集和核酸提取将A2接种至PDA上培养收集菌丝。gDNA 的提取按照OMEGA HP Fungal DNA Kit 说明书操作。RNA 的提取按照OMEGA Fungal RNA Kit说明书操作。
1.2.2Lac2基因的PCR扩增采用同源克隆策略。经本地查找橡胶树胶孢炭疽菌全基因组测序数据,设计特异引物对Lac2-F(5′-ATGGTCGCCATCAAAGACCTTATGAA-3′)和Lac2-R(5′-TTACTGACCGCTGTCAATGAC-3′)扩增漆酶基因,以C. gloeosporioides基因组gDNA为模板,扩增体系(50μL):10×PCR Buffer 5μL;dNTP Mixture(各2.5 mmol/L)4 μL;Lac2-F(10 μmol/L)和Lac2-R(10 μmol/L)各2 μL;TaKaRa Taq HS 0.5μL;DNA模板1μL;加ddH2O补至50 μL。扩增程序:95 ℃ 5 min;95 ℃ 50 s,60 ℃ 40 s,72 ℃ 2min,35个循环;72 ℃ 10 min。PCR产物的凝胶回收、连接分别按照OMEGA和TaKaRa相应试剂盒操作。用载体通用引物M13-47和RV-M进行PCR 鉴定后,选择阳性克隆测序(华大基因生物有限公司)。
1.2.3Lac2基因的RT-PCR扩增用DNAMAN 1.0软件对所得Lac2基因全序列与GenBank中的嗜热毁丝霉(Myceliophthora thermophila)lac基因cDNA序列(索取号:XM-003659795.1)比对分析后,用推定的cDNA序列设计引物对RLac2 -F(5′-TCGCCATCAAAGACCTTATGAAA-3′)和RLac2 -R(5′-ACCTGGTCAACCTTGGACGGGAT-3′)用于扩增Lac2的cDNA。RT-PCR按照PrimeScriptTM Ⅱ High Fidelity RT-PCR Kit说明书操作,其余同上。
1.2.4Lac2序列分析用Expasy 的ProtParam 在线分析软件(http://web.expasy.org/protparam/)分析该蛋白序列组分;利用Clustal X 1.81 软件进行多重序列比对,采用MEGA 5.1 软件通过neighbor-joining 方法构建系统进化树,其中bootstrap设为1000 replicates。其余参照文献[12]。
1.2.5Lac2基因在分生孢子不同萌发时段的半定量表达分析通过筛选,选用真菌18S rRNA通用引物作为内参基因(SR5:5′-GTGCCCTTCCGTCAATT-3′,SR7R:5′-AGTTAAAAAGCTCGTAGTTG-3′);根据已获得的Lac2 cDNA序列,设计半定量引物对qLac2 -F(5′-ACAACGGTAGCGTTCGTGTTATTG-3′)和qLac2 -R(5′-ACCTGGTCAACCTTGGACGGGAT-3′)。参照文献[13],配制1×107/mL的孢子悬浮液,取6 mL均匀涂布于保鲜膜上,保湿密闭,置于28 ℃ 黑暗培养,分别收集处理0、1、2、4、6、9、12、18 h 后的分生孢子,提取总RNA。扩增体系(25 μL):Primer STAR HS 12.5 μL;qLac2 -F(10 μmol/L)和qLac2 -R(10 μmol/L)各1 μL;cDNA模板1μL;加ddH2O补至25 μL。扩增程序:95 ℃ 3 min;95 ℃ 30 s,65 ℃ 40 s,72 ℃ 40 s,34个循环;72 ℃ 7 min。
2结果与分析
2.1芒果胶胞炭疽菌Lac2基因的克隆
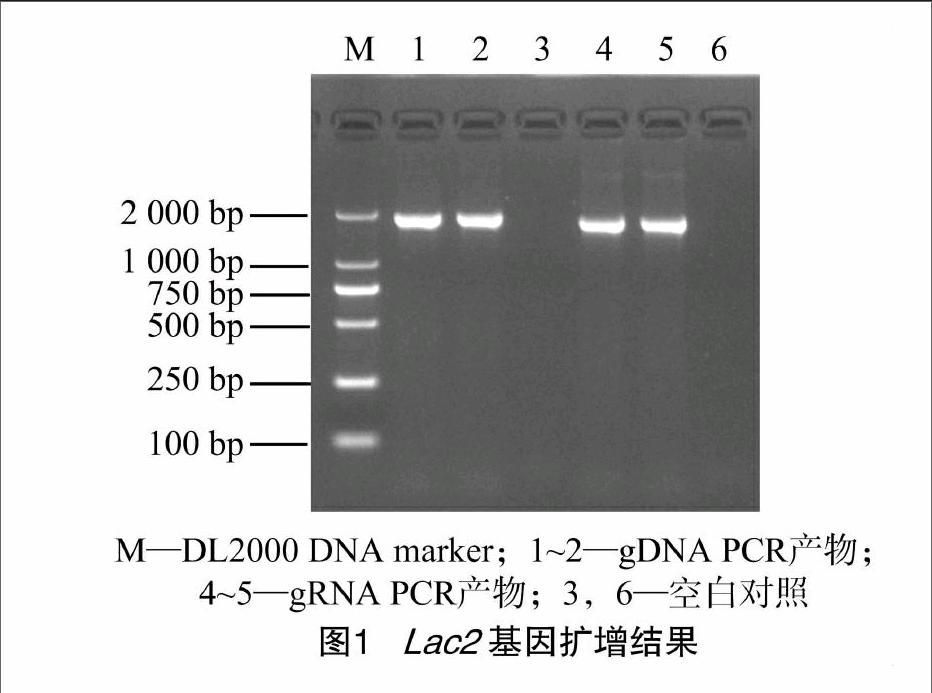
用设计的特异引物对Lac2 -F和Lac2 -R,以芒果胶胞炭疽菌gDNA为模板扩增获得了约2 000 bp的片段(图1),与预期的序列大小一致。双向测通结果表明,插入的目的片段大小为1 966 bp。经BLASTN分析发现,该片段与多个不同物种的漆酶基因序列有很高的相似性,最高达81%,但与已报道的芒果胶胞炭疽菌Lac1[12]的同源性仅为24.58%,说明该片段为芒果胶胞炭疽菌的另一个漆酶基因,命名为Lac2基因。用Primer 5.0软件推测Lac2基因编码的氨基酸序列,发现目的片段含完整的开放阅读框。
2.2RT-PCR扩增获得Lac2的cDNA序列
经RT-PCR扩增获得约1 800 bp的cDNA片段(图1),测序表明大小为1 785 bp。该序列已提交GenBank,登录号为KF924625。
2.3Lac2序列分析
2.3.1Lac2基因全长DNA与cDNA序列分析比对分析Lac2基因DNA和cDNA序列,发现含1个1 785 bp大小的开放阅读框,起始密码子ATG位于第1~3 bp处,终止密码子TAA位于第1 964~1 966 bp处,自ATG到TAG的DNA全长1 966 bp,其中存在3个大小分别57、71、53bp的内含子(位于第86~143、565~636和1 564~1 617碱基处),是典型的真菌内含子的长度(49 ~85 bp),所有内含子均符合GT-AG法则。用BioEdit软件分析结果表明:Lac2基因编码序列的双链核苷酸分子量大小为1 087.22 ku,(G+C)摩尔百分含量为 56.86%,(A+T)摩尔百分含量为43.14%,可见该编码区富含GC碱基,其中C的摩尔百分含量最高,为34.01%。
2.3.2Lac2基因预测蛋白的生化特征分析用Primer 5.0软件预测Lac2基因编码594个氨基酸,其中参照文献[14]发现其中含有5个可能的N-糖基化位点(Asn-X-Thr/Ser)(分别位于第9~11、152~154、394~396、401~403、580~582氨基酸处),分子量约为65.53 ku,包括59个强碱性氨基酸(KRH)、47个强酸性氨基酸(DE)、242个非极性氨基酸(AVLIPFWM)、352个极性氨基酸(GSTCYNQKRHDE),等电点(pI)为6.56,可见是一种偏酸性蛋白质,富含极性氨基酸,特别是天冬酰胺(Asn)、甘氨酸(Gly)的含量分别达9.1%和8.2%。在真菌漆酶中,由于Cu原子结合保守区的Cys残基下游的第10个氨基酸残基种类对漆酶氧化还原潜能具有重要作用,Shin据此将漆酶分为3类:1型(Met)、2型(Leu)和3型(Phe)[15],按此分类标准,Lac2属于2型漆酶。漆酶属含铜的多酚氧化酶,对Lac2的氨基酸序列分析发现它具有4个铜离子结合区域(His-X-His)、10个铜离子结合位点(图2)。
2.3.3Lac2基因预测蛋白的同源性分析将该基因编码的氨基酸序列提交NCBI进行BLASTP同源比对,分析表明该漆酶与已发表的C. orbiculare MAFF 240422 laccase-1 precursor(ENH77191.1)、C. higginsianum multicopper oxidase(CCF34868.1)、C. graminicola M1.001 multicopper oxidase(EFQ36357.1)、Trichoderma sp. T01 laccase(ACS45199.1)、Gaeumannomyces graminis var. tritici R3-111a-1 laccase-1(EJT70878.1)、Fusarium oxysporum f.sp. cubense race 1 Laccase-2(ENH66075.1)、Metarhizium acridum CQMa 102 Lcc1(EFY87101.1)、F. proliferatum laccase(AAK72901.1)和F.oxysporum f.sp. lycopersici Lcc1(ABS19938.1)基因编码的蛋白同源性较高,相似性分别为79%、78%、70%、63%、61%、61%、61%、61%和60%。系统聚类结果表明(图3):Lac2预测蛋白可能与西瓜炭疽菌(C. orbiculare)的laccase类或禾生炭疽菌(C. graminicola)的multicopper oxidase聚为一类。
2.3.4Lac2基因预测蛋白的保守结构域分析利用NCBI 的Blast 软件(http://www.nebi.nlm.nih.gov/strueture/cdd/wrpsb.cgi)进行Lac2基因预测蛋白的结构域分析(图4),发现该蛋白中含有Cu-oxidase 保守结构域,说明Lac2蛋白属于含铜的氧化物酶类。
2.4Lac2基因在分生孢子不同萌发时段的半定量表达分析
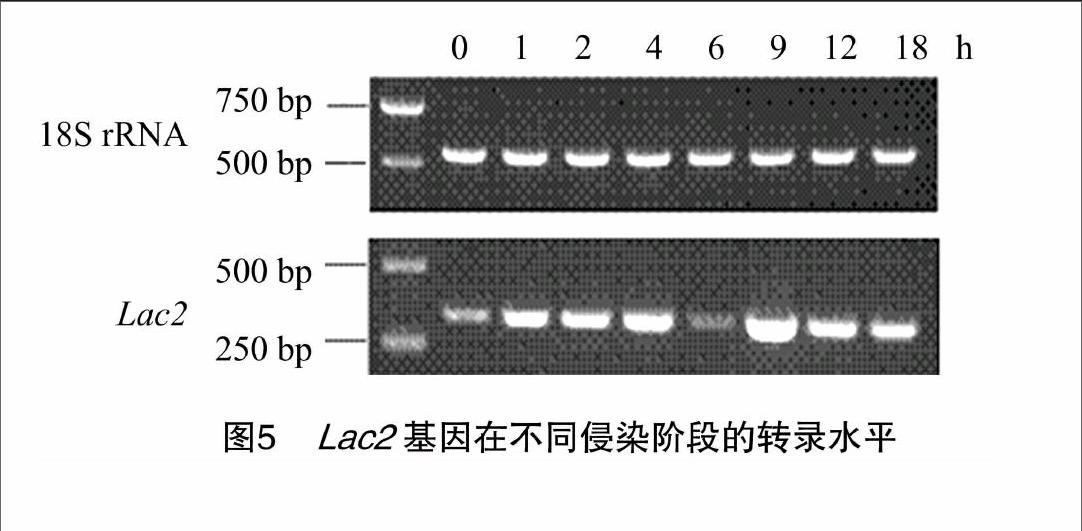
在胶孢炭疽菌分生孢子萌发过程中,Lac2 在不同时期表达量不尽相同(图5),有明显的升降趋势,总体呈“升—降—升—降”趋势,0 h Lac2 的表达比较弱,0~4 h表达量逐渐上升,至4 h 达到第1个波峰,而6 h时突然减弱,9 h 又突然达到第2个波峰,随后逐渐减弱;总体上在处理6 h 后表达量最低,9 h 后达到最高。据此初步推测Lac2与C. gloeosporioides分生孢子萌发后侵染寄主有关。
3讨论
漆酶一般含有4个铜离子结合区,不同漆酶基因所编码的氨基酸序列的同源性较抵,但铜原子结合区保守性高,该区是鉴定获得基因是否为漆酶基因的主要依据[7]。笔者所在实验室先后发现的胶孢炭疽菌的2个漆酶基因Lac1和Lac2的氨基酸序列同源性仅24.58%,而对应的铜原子结合区保守性较高。可见,根据铜原子结合区保守序列设计简并引物是克隆漆酶基因有效途径。
漆酶基因广泛存在于子囊菌和担子菌中,在不同的生长条件、生长周期及侵染过程的表达量不同。在不同培养条件下,小麦全蚀病菌(Gaeumannomyces graminis var. tritici)的3种同工漆酶的表达量存在明显差别,其中Lac1均表达,Lac2只在含有CuSO4的培养基上表达,而Lac3的表达仅在含寄主滤液的基础培养基上才能检测到[16];番茄枯萎病菌(F. oxysporum f. sp. lycopersici)的Lac2、Lac4和Lac5在整个侵染过程中均不表达,而Lac1、Lac3和Lac9能表达,但时间上存在差异[3]。C. gloeosporioides孢子悬浮液处理9 h后,80%以上的炭疽菌分生孢子萌发并形成附着胞,开始侵染寄主[17];本试验Lac2在侵染0~18 h时段中,9 h表达量最高,因此推测Lac2可能与C. gloeosporioides分生孢子萌发侵染寄主相关。
在不同植物病原真菌中的漆酶的功能差异较大。番茄枯萎病菌(F. oxysporum f. sp. lycopersici)的3个漆酶基因(lac1、lac3和lac5)的缺失突变体对致病力均无影响,而Δlac3与氧化胁迫能力及对酚类化合物的敏感性相关,Δlac1与氧化胁迫能力及漆酶酶活相关[3];玉米大斑病菌(Setosphaeria turcica)的漆酶基因(StLAC1)缺失突变体不能形成分生孢子,附着胞膨压、致病力、黑色素合成明显减弱,菌落颜色由灰黑色变为灰白色[4];与本研究克隆的Lac2基因编码的蛋白同源性达70%以上的C. orbiculare laccase-1 precursor(ENH77191.1)、C.higginsianum multicopper oxidase(CCF34868.1)和C.graminicola multicopper oxidase(EFQ36357.1)未见相应基因功能鉴定的报道,而Lac2编码蛋白与F. oxysporum f. sp. lycopersici的Lac1编码的漆酶有较高的序列相似性,且Lac1与抗氧化反应及漆酶酶活有关,因此推测芒果胶胞炭疽菌的Lac2基因可能也有相似的功能。但由于芒果胶胞炭疽菌与上述真菌在生活史、侵入寄主的方式、致病因子等方面均存在差异,该基因的具体功能还有待进一步研究。
参考文献:
[1]靳蓉,张飞龙. 漆酶的应用技术[J]. 中国生漆,2013,32(2):35-42.
[2]Widsten P,Kandelbauer A. Laccase applications in the forest products industry:a review[J]. Enzyme and Microbial Technology,2008,42(4):293-307.
[3]Cordoba C D,Roncero M I. Functional analyses of laccase genes from Fusarium oxysporum[J]. Phytopathology,2008,98(5):509-518.
[4]詹旭. 玉米大斑病菌漆酶基因StLAC1在黑色素合成中的功能分析[D]. 保定:河北农业大学,2011.
[5]Fan X,Zhou Y,Xiao Y,et al. Cloning,expression and phylogenetic analysis of a divergent laccase multigene family in Auricularia auricula-judae[J]. Microbiological Research,2014,169(5/6):453-462.
[6]肖书笑,田呈明,王永林. 黄栌枯萎病菌漆酶基因的鉴定及其在微菌核形成过程中的表达研究[J/OL]. [2013-12-20]. http://www.paper.edu.cn/releasepaper/content/201312-625.
[7]Tsuji G,Fujikawa J,Ishida H. Laccase gene LAC1 of Colletotrichum lagenarium is not essential for melanin biosynthesis and pathogenicity[J]. Journal of General Plant Pathology,2001,67(3):182-190.
[8]Lin S Y,Okuda S,Ikeda K,et al. Lac2 encoding a secreted laccase is involved in appressorial melanization and conidial pigmentation in Colletotrichum orbiculare[J]. Molecular Plant-Microbe Interactions,2012,25(12):1552-1561.
[9]王葵娣,王文华,郑服丛. 炭疽菌附着胞的研究进展[J]. 中国农学通报,2007,23(1):265-270.
[10]王洪凯,林福呈,王政逸. 植物病原真菌附着胞的机械穿透力[J]. 菌物学报,2004,23(01):151-157.
[11]Butler M J,Gardiner R B,Day A W. Melanin synthesis by Sclerotinia sclerotiorum[J]. Mycologia,2009,101(3):296-304.
[12]韦运谢,刘晓妹,张贺,等. 芒果炭疽病菌漆酶基因lac1的克隆与序列特征分析[J]. 果树学报,2013,30(2):202-206.
[13]Takano Y,Kubo Y,Kuroda I,et al. Temporal transcriptional pattern of three melanin biosynthesis genes,PKS1,SCD1,and THR1,in appressorium-differentiating and nondifferentiating conidia of Colletotrichum lagenarium[J]. Applied and Environmental Microbiology,1997,63(1):351-354.
[14]李剑凤,洪宇植,肖亚中. 栓菌420漆酶同工酶B基因克隆及异源表达[J]. 生物学杂志,2007,24(3):25-28.
[15]Shin K S,Lee Y J. Purification and characterization of a new member of the laccase family from the white-rot basidiomycete Coriolus hirsutus[J]. Archives of Biochemistry and Biophysics,2000,384(1):109-115.
[16]Litvintseva A P,Henson J M. Cloning,characterization,and transcription of three laccase genes from Gaeumannomyces graminis var. tritici,the take-all fungus[J]. Applied and Environmental Microbiology,2002,68(3):1305-1311.
[17]王科. 黄柏炭疽病(Colletotriehum gloeosporioides)的研究[D]. 雅安:四川农业大学,2010.
猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2020年24期)2020-12-15
小学科学(学生版)(2019年5期)2019-05-21
兽医导刊(2016年12期)2016-05-17
现代检验医学杂志(2015年4期)2015-02-06
