
人自噬相关基因12真核表达载体的构建及表达
2015-11-29 08:32纪贝贝张鹏徐小洁梁迎春黄蓉范忠义李玲郭靖洪甜叶棋浓杜楠
生物技术通讯 2015年6期
纪贝贝 ,张鹏,徐小洁,梁迎春,黄蓉,范忠义,李玲,郭靖,洪甜,,叶棋浓,杜楠
1.军事医学科学院 生物工程研究所,北京 100850;2.解放军总医院 第一附属医院,北京 100048;3.解放军302医院 普外诊疗中心,北京 100039
人自噬相关基因12(autophagy related gene 12,ATG12)是自噬相关基因家族成员,对自噬的形成至关重要[1]。自噬是细胞维持稳态的一种机制,细胞内多数衰老长半衰期蛋白质可通过自噬途径降解,从而维持细胞内稳定[2]。研究发现自噬参与肿瘤、病原体免疫的过程,自噬可通过一系列机制将肿瘤细胞[3]、入侵的病原体[4]等降解,其机制为,来自高尔基复合体或粗面内质网的单层或双层膜将抗原包裹后形成自噬泡,自噬泡的外膜与溶酶体膜融合,内膜及其包裹的物质进入溶酶体腔,被溶酶体中的酶水解,从而维持机体稳定。
ATG12 可以和ATG5、ATG16 结合形成复合物,该复合物参与自噬体的形成[5],在自噬过程中发挥重要作用[6-7]。ATG3 蛋白是一种泛素样缀合酶,它和ATG10 是促进ATG8 和ATG12 泛素样蛋白结合的2种E2 样酶[8]。有文献报道称,在自噬过程中ATG12可与ATG3 相互作用[9]。鉴于ATG12 在自噬中的重要作用,我们拟构建ATG12 的真核表达载体,并验证表达的myc-ATG12 融合蛋白与ATG3 的相互作用,为进一步研究ATG12与自噬的关系奠定基础。
1 材料和方法
1.1 材料
人胚肾293T细胞由本室传代培养;pXJ-40-myc载体为本实验室保存;Flag-ATG3 质粒为本实验室前期构建;VigoFect 为威格拉斯生物技术有限公司产品;限制性内切酶、DNA 连接酶、PCR 试剂均购自TaKaRa 公司;上、下游引物由北京赛百盛生物技术有限公司合成;质粒提取、胶回收、PCR 回收试剂盒购自Promega公司;HRP标记的抗myc标签鼠单克隆抗体(myc-HRP)和抗Flag标签鼠单克隆抗体(Flag-HRP)购自Sigma公司;DMEM及小牛血清购自Gibco公司;测序由北京奥科生物技术有限责任公司完成。
1.2 myc-ATG12重组质粒的构建与测序
以本实验室保存的人乳腺文库为模板,根据文献报道[10]的ATG12 编码序列合成上游引物(5'-CG GGATCCATGGCGGAGGAGCCGCAGTCTG-3')和下游引物(5'-CCGCTCGAGTCATCCCCACGCCTGAG ACTTGC-3'),PCR 扩增人ATG12 的编码序列(扩增条件:95℃预变性5 min;95℃变性30 s,60℃退火30 s,72℃延伸30 s,30个循环;72℃延长7 min),用胶回收试剂盒回收PCR产物。
用BamHⅠ/XhoⅠ双酶切pXJ-40-myc 载体,经10 g/L 琼脂糖凝胶电泳后,胶回收载体大片段;将PCR片段回收后再用BamHⅠ/XhoⅠ酶切,形成带有粘端的双链,用T4DNA 连接酶连接入pXJ-40-myc载体;转化大肠杆菌DH5α,挑选克隆,振荡培养并提质粒,用BamHⅠ/XhoⅠ双酶切鉴定,酶切鉴定正确的克隆送北京奥科生物公司测序。
1.3 哺乳动物细胞转染和Western印迹检测
按常规方法进行转染,用不含双抗、含100 mL/L 胎牛血清的DMEM 培养基将293T 细胞接种于6 cm 皿中,接种量以转染时细胞密度达80%为宜,培养24 h 后进行转染,转染前1 h 换液。将4 μL VigoFect与200 μL NaCl混合,再将总量为10 μg的重组质粒与200 μL NaCl 混合,然后将上述2 种溶液轻轻混合,室温放置15 min,加入6 cm皿中,并以同样方法转染空pXJ-40-myc 载体作为对照,37℃、5% CO2常规培养,4~6 h换液。质粒转染293T细胞24 h 后收集细胞蛋白,加入2×SDS 加样缓冲液,煮沸10 min,高速离心2 min,取上清液进行SDSPAGE后电转移至硝酸纤维素膜上;用5%脱脂奶粉于4℃封闭过夜,加入用5%脱脂奶粉以1∶5000稀释的用HRP 标记的抗myc 标签鼠单克隆抗体,室温轻摇1 h,TBST洗膜3次,每次5 min;用化学发光法显色5 min,压片显影。
1.4 免疫共沉淀检测ATG12和ATG3的相互作用
将人胚肾293T 细胞接种于6 cm 皿中,用重组myc-ATG12 与Flag-ATG3 共转染细胞,转染后4~6 h 换液,常规培养24 h 后收集细胞,加入中盐(0.25 mol/L)IP 缓冲液后与myc-beads 于4℃结合4~6 h,3000 r/min 离心5 min,经中盐IP 缓冲液再次漂洗后,加入与myc-beads 等量的2×SDS 加样缓冲液,煮沸10 min,12 000 r/min 离心2 min,取上清液进行SDS-PAGE后电转移至硝酸纤维素膜上,用5%脱脂奶粉于4℃封闭1 h,加入用5%脱脂奶粉以1∶5000稀释的用HRP标记的抗myc和Flag标签鼠单克隆抗体,室温轻摇1 h,TBST 洗膜3 次,每次5 min,用化学发光法显色,5 min后压片显影。
2 结果
2.1 myc-ATG12重组质粒的构建与鉴定
以实验室保存的人乳腺文库为模板,PCR 扩增人ATG12 的编码序列,获得约420 bp 的DNA 片段,与预期一致(图1)。将PCR 产物用BamHⅠ/XhoⅠ双酶切后,与经同样双酶切的pXJ-40-myc 载体连接,转化大肠杆菌DH5α,挑选阳性克隆提质粒,经酶切鉴定,可切出2条长度分别约为5000和420 bp的条带,而相应的空载体酶切只见大片段,符合预期结果(图2)。测序结果表明,插入片段的DNA 序列与人ATG12基因的编码序列完全一致(数据略)。

图1 PCR扩增人ATG12的编码序列

图2 重组质粒myc-ATG12的BamHⅠ/XhoⅠ双酶切电泳图谱
2.2 Western 印迹检测myc-ATG12 在293T 细胞中的表达
将构建的myc-ATG12 重组质粒和空载体分别转染293T 细胞,24 h 后提取蛋白进行SDS-PAGE,Western 印迹检测myc-ATG12 蛋白的表达。结果显示,转染重组质粒后,用myc-HRP 抗体能够在相对分子质量约17×103处检测到明显的特异性条带,而空载体则无条带(图3)。说明myc-ATG12重组蛋白在293T细胞中能够正确表达。
2.3 免疫共沉淀检测ATG12与ATG3的相互作用
根据文献报道,ATG12 可与ATG3 蛋白相互作用。为进一步证实构建的myc-ATG12 重组质粒正确,且能够表达正确的融合蛋白,将重组myc-ATG12 质粒与Flag-ATG3 质粒共转染293T 细胞,24 h常规培养后收集蛋白,免疫共沉淀分析观察显示,myc-ATG12 融合蛋白与Flag-ATG3 蛋白具有相互作用,而myc 空载体在同一位置无此条带,表明ATG12 与ATG3 蛋白能够在体内特异地相互作用,而myc 标签不影响ATG12 的结构及其生物学功能(图4)。进一步证明构建的重组myc-ATG12质粒正确,且能够正常表达。
3 讨论
自噬是一种普遍而又重要的生命现象,广泛参与多种生理和病理过程,特别是与肿瘤发生密切相关。目前,自噬的调控作用被广泛关注并加以研究,以求从分子水平上对肿瘤进行治疗,从而达到根治的目的[11-13]。针对ATG的调控是现在自噬研究的热点。根据其在自噬过程的不同阶段发挥作用的不同,可将ATG 分为以下三类:一是ATG1-ATG11-ATG17-ATG20-ATG24-ATG29-ATG31 和ATG13-ATG8 复合体,二是ATG6-ATG14-Vps34-Vps15 复合体,三是ATG12-ATG5-ATG16和LC3-II-PE泛素样蛋白系统[14]。可见ATG12与自噬体的形成密切相关。有研究显示ATGl2缺失的小鼠胚胎干细胞前自噬体结构和自噬体的数量明显减少[15]。
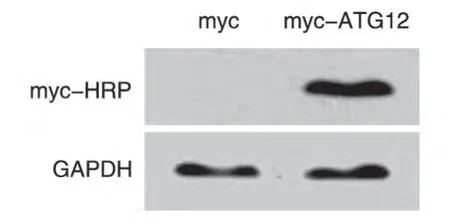
图3 Western印迹检测myc-ATG12的表达

图4 免疫共沉淀分析验证重组ATG12与ATG3在蛋白水平上具有相互作用
ATG12 定位于细胞质和自噬体中,同时介导自噬和凋亡两条通路[16],在自噬过程中和ATG5的结合是不可逆的,同时与ATG3 的结合区域也有很多疑惑待解决[17]。本实验构建的myc-ATG12在真核细胞中获得了表达,且表达的融合蛋白能够与ATG3 蛋白相互作用。ATG12 在真核细胞中的成功表达,是继续深入研究其在自噬中的作用机制,以及探讨其利用价值的基础。
[1]Mariño G,López-Otín C.Autophagy:molecular mechanisms,physiological functions and relevance in human pathology[J].Cell Mol Life Sci,2004,61(12):1439-1454.
[2]Reggiori F,Klionsky D J.Autophagy in the eukaryotic cell[J].Eukaryot Cell,2002,1(1):11-21.
[3]Tóth S,Nagy K,Pálfia Z,et al.Cellular autophagic capacity changes during azaserine-induced tumour progression in the rat pancreas.Up-regulation in all premalignant stages and down-regulation with loss of cycloheximide sensitivity of segregation along with malignant transformation[J].Cell Tissue Res,2002,309(3):409-416.
[4]Rich K A,Burkett C,Webster P.Cytoplasmic bacteria can be targets for autophagy[J].Cell Microbiol,2003,5(7):455-468.
[5]Suzuki K,Kirisako T,Kamada Y,et al.The pre-autophagosomal structure organized by concerted functions of APG genes is essential for autophagosome formation[J].EMBO J,2001,20(21):5971-5981.
[6]Thompson A R,Doelling J H,Suttangkakul A,et al.Autophagic nutrient recycling in Arabidopsis directed by the ATG8 and ATG12 conjugation pathways[J].Plant Physiol,2005,138(4):2097-2110.
[7]Suzuki N N,Yoshimoto K,Fujioka Y,et al.The crystal structure of plant ATG12 and its biological implication in autophagy[J].Autophagy,2005,1(2):119-126.
[8]Flanagan M D,Whitehall S K,Morgan B A.An Atg10-like E2 enzyme is essential for cell cycle progression but not autophagy in Schizosaccharomyces pombe[J].Cell Cycle,2013,12(2):271-277.
[9]Radoshevich L,Murrow L,Chen N,et al.ATG12 conjugation to ATG3 regulates mitochondrial homeostasis and cell death[J].Cell,2010,142(4):590-600.
[10]Umemiya-Shirafuji R,Galay R L,Maeda H,et al.Expression analysis of autophagy-related genes in the hard tick Haemaphysalis longicornis[J].Vet Parasitol,2014,201(1-2):169-175.
[11]Ding Z B,Shi Y H,Zhou J,et al.Association of autophagy defect with a malignant phenotype and poor prognosis of hepatocellular carcinoma[J].Cancer Res,2008,68(22):9167-9175.
[12]Wu G,Li H,Ji Z,et al.Inhibition of autophagy by autophagic inhibitors enhances apoptosis induced by bortezomib in non-small cell lung cancer cells[J].Biotechnol Lett,2014,36(6):1171-1178.
[13]Cufí S,Vazquez-Martin A,Oliveras-Ferraros C,et al.Autophagy-related gene 12(ATG12) is a novel determinant of primary resistance to HER2-targeted therapies:utility of transcriptome analysis of the autophagy interactome to guide breast cancer treatment[J].Oncotarget,2012,3(12):1600-1614.
[14]Geng J,Klionsky D J.The Atg8 and Atg12 ubiquitin-like conjugation systems in macroautophagy.'Protein modifications:beyond the usual suspects' review series[J].EMBO Rep,2008,9(9):859-864.
[15]Mizushima N,Yamamoto A,Hatano M,et al.Dissection of autophagosome formation using Apg5-deficient mouse embryonic stem cells[J].J Cell Biol,2001,152(4):657-668.
[16]Rubinstein A D,Eisenstein M,Ber Y,et al.The autophagy protein Atg12 associates with antiapoptotic Bcl-2 family members to promote mitochondrial apoptosis[J].Mol Cell,2011,44(5):698-709.
[17]Metlagel Z,Otomo C,Takaesu G,et al.Structural basis of ATG3 recognition by the autophagic ubiquitin-like protein ATG12[J].Proc Natl Acad Sci USA,2013,110(47):18844-18849.
猜你喜欢
河北北方学院学报(自然科学版)(2022年11期)2022-02-03
中国科技纵横(2021年24期)2021-03-02
中国乳业(2019年12期)2020-01-17
中国食品(2019年1期)2019-09-10
中成药(2018年6期)2018-07-11
食品科学(2018年10期)2018-05-23
中国乳业(2018年5期)2018-01-26
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
西南军医(2015年6期)2015-01-23
