
马铃薯立枯丝核菌生物学特性研究
2015-02-14 06:14台莲梅赵巧兰靳学慧金光辉
植物保护 2015年1期
台莲梅, 赵巧兰, 靳学慧, 金光辉
(黑龙江八一农垦大学农学院,大庆 163319)
马铃薯立枯丝核菌生物学特性研究
台莲梅, 赵巧兰, 靳学慧, 金光辉
(黑龙江八一农垦大学农学院,大庆 163319)
采用生长速率法研究了黑龙江省马铃薯立枯丝核菌(Rhizoctonia solaniKühn)菌丝生长的条件。结果表明病菌菌丝在10~35℃均能生长,适宜温度是25~30℃,30℃菌丝生长最快。在p H为3~12的范围内病菌菌丝均能生长,以p H 5~7菌丝生长速度较快。黑暗对菌丝生长有促进作用,光照可促进菌核的形成。不同碳源、氮源对立枯丝核菌菌丝生长的影响均较明显,可溶性淀粉和酵母粉对菌丝生长有显著的促进作用。立枯丝核菌菌丝体及菌核的致死温度分别为49℃和51℃。明确了黑龙江省马铃薯立枯丝核菌生长的环境条件,可为病害防治提供理论依据。
马铃薯; 立枯丝核菌; 生物学特性
马铃薯立枯丝核菌病,又称黑痣病、茎基腐病等,是土传性病害。在我国,立枯丝核菌病最早是1922年和1932年分别于台湾和广东发现,目前它的分布已经相当普遍[1]。近年来,随着马铃薯产业的迅速发展,种植面积不断扩大,加上其他农作物种植结构的调整,导致马铃薯不能正常轮作,马铃薯立枯丝核菌病有加重的趋势。植株发病后,不仅薯块变小,产量降低,而且品质变劣,重病区可减产51%[2]。丝核菌侵染幼芽后,有的幼芽在出土前就腐烂死亡,形成芽腐,造成田间缺苗;出土后染病植株茎基形成褐色凹陷病斑,受害重的可造成立枯、顶端萎蔫或叶片卷曲等症状。薯块受害后,表面有许多形状、大小不规则的黑色菌核。菌丝生长温度最低为4℃,最适为23℃,生长上限温度为32~33℃之间,34℃时停止生长[2]。根据菌丝是否融合,将立枯丝核菌划分为13个融合群(AG1~AG13)[3-4]。融合亚群已有18个[5]。Scholte等[6]认为AG-3能够引起马铃薯丝核菌病的发生,其寄主范围很小。Virgen等[7]根据来自墨西哥15个地点的取样调查得出,AG-3和AG-4的发生率分别为73.5%和26.5%。其他融合群如AG1、AG2、AG5、AG7和AG9在马铃薯黑痣病中也有发现[8]。马铃薯立枯丝核菌病的防治,目前主要采用化学药剂进行拌种、沟施[9-10]。关于该病其他方面的研究报道较少。本试验对采集的马铃薯丝核菌茎溃疡病样进行病原菌分离、鉴定,对病菌的生物学特性进行研究,以期为病害发生规律的研究和病害综合防治提供科学依据。
1 材料与方法
1.1 供试菌种
供试病菌:分离自黑龙江农垦总局九三管理局植保站试验田的马铃薯病样。28℃条件下,PDA上培养2 d,取直径为6 mm的等龄菌饼供试。
1.2 不同温度对病菌菌丝生长的影响
分别设置5、10、15、20、25、30、35℃,共7个处理。每个处理4次重复。将菌饼接种于PDA平板上,36 h后用十字交叉法测量菌落直径。
1.3 不同p H对病菌菌丝生长的影响
PDA培养基灭菌以后,无菌条件下用1 mol/L NaOH和1 mol/L HCl溶液调节PDA培养基的p H,分别设置3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0、12.0共10个p H梯度。每个处理4次重复。将菌饼接种于PDA平板上,于28℃恒温培养箱中培养。36 h后用十字交叉法测量菌落直径。
1.4 不同碳源对病菌菌丝生长的影响
供试碳源:葡萄糖、半乳糖、乳糖、可溶性淀粉、麦芽糖、果糖,将查氏培养基中的蔗糖置换其他碳源,并且以无糖为对照配成不同碳源的培养基,共7个处理,每处理4次重复。
查氏培养基:2.00 g KNO3,1.00 g KH2PO4,0.5 g KCl,0.01 g FeSO4,0.50 g MgSO4·7H2O,20.00 g琼脂,30.00 g蔗糖,1 000 g蒸馏水。将菌饼接种于不同碳源培养基中,以不加碳源为对照,28℃恒温培养箱中培养。36 h后测量菌落直径。
1.5 不同氮源对病菌菌丝生长的影响
供试氮源:蛋白胨、半胱氨酸、硝酸钠、酵母粉、丙氨酸、尿素;查氏培养基中的硝酸钾置换成其他氮源,并设无氮对照,配成不同氮源的培养基,共7个处理,每处理4次重复。将菌饼接种于不同氮源培养基中。28℃恒温培养箱中培养36 h后,测量菌落直径。
1.6 光照对病菌菌丝生长及菌核形成的影响
分别设置连续光照、连续黑暗、12 h光暗交替3个处理,每个处理4次重复。将菌饼接种于PDA平板上。28℃恒温培养,36 h后测量菌落直径。观察各处理最初形成菌核天数,以及10 d后形成菌核的数量。
1.7 病菌菌丝及菌核致死温度的测定
菌丝体致死温度测定:将6 mm菌饼置于同样大小的无菌试管中,加入2 mL的无菌水。分别放入40、45、46、47、48、49、50、55、60℃不同温度恒温水浴锅中加热10 min,后用冷水将试管迅速冷却。然后将试管中的菌饼取出放于PDA平板上,未经温水处理的菌饼为对照,28℃条件下培养36 h后,记录菌丝的生长情况。每处理4次重复。
菌核致死温度测定:利用PDA培养基上生长1个月的成熟菌核为材料,将菌核放入无菌试管中,在40、45、50、51、52、53、54℃不同温度恒温水浴锅中加热10 min,并用冷水将试管迅速冷却。将菌核取出放置于PDA平板上,未经温水处理的菌核为对照。每个处理4次重复。28℃下进行光照培养36 h后,观察菌核萌发情况。
2 结果与分析
2.1 温度对病菌菌丝生长的影响
在PDA培养基上,病菌菌丝在5℃时不生长,在10~35℃均能生长,但在25~30℃生长较快,30℃生长最快。25、30℃菌落直径分别为76.4 mm和81.6 mm。温度35℃时,病菌菌丝生长速度明显降低(图1)。
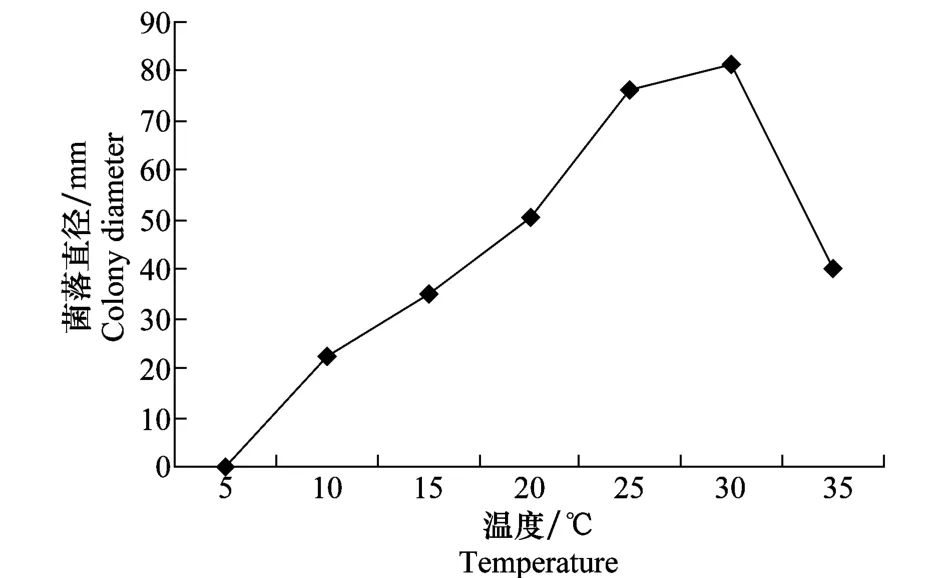
图1 温度对立枯丝核菌菌丝生长的影响Fig.1 Effects of temperature on mycelial growth ofR.solani
2.2 p H对病菌菌丝生长的影响
不同的p H条件对菌丝生长影响差异明显。p H在3~12范围内病菌的菌丝均能生长,而p H 5~7菌丝的生长速度相对较快,36 h菌落直径可以达到63.1~73.0 mm,p H为7时,菌丝生长速度最快(图2)。另外,从试验结果中还可以看出,病菌在酸性条件下有利于生长。
2.3 碳源对病菌菌丝生长的影响
病菌在供试的7种碳源中均可生长(表1)。其中以可溶性淀粉为碳源的菌丝生长最快,极显著高于其他碳源(P<0.01),其次是蔗糖为碳源的培养基中菌丝生长较快,显著高于其他碳源(P<0.05),在果糖培养基中菌丝生长效果差。
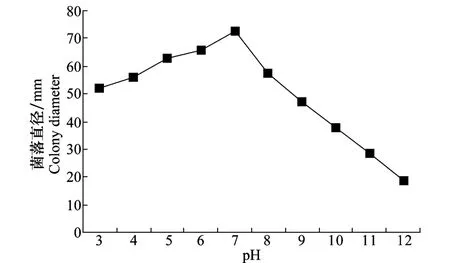
图2 pH对立枯丝核菌菌丝生长的影响Fig.2 Effects of pH on mycelial growth ofR.solani
2.4 氮源对病菌菌丝生长的影响
病菌在供试的7种氮源中均可生长(表1)。其中以酵母粉为氮源生长最好,其次是蛋白胨为氮源生长较好,而对半胱氨酸的利用最差,各处理间差异极显著。
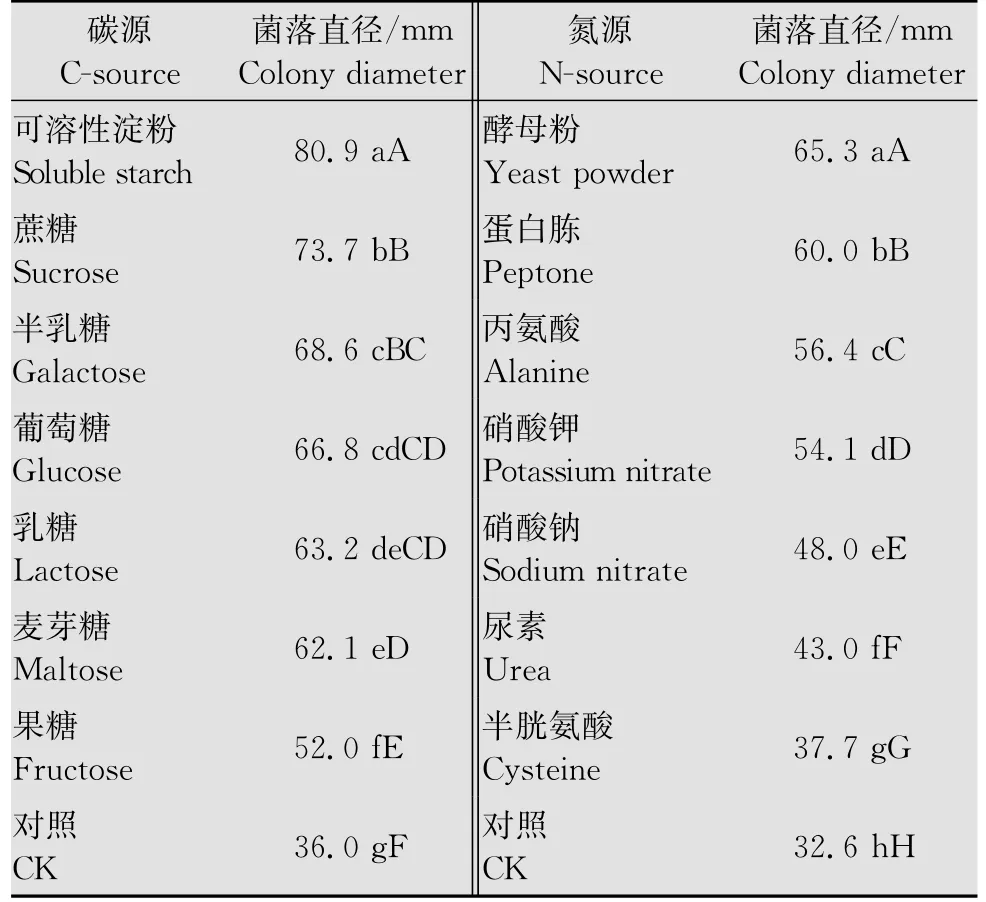
表1 不同碳、氮源培养基培养的立枯丝核菌菌落直径1)Table 1 Colony diameters ofR.solaniin different carbon and nitrogen sources
2.5 光照对病菌菌丝生长及菌核形成的影响
对病菌菌丝体及菌核进行光照、黑暗、光暗交替处理,结果见表2。在全黑暗情况下菌丝生长最快,菌落平均直径为80 mm;其次是光暗交替,菌落直径为74 mm,2个处理之间差异不显著,但与全光照处理差异极显著。

表2 不同光照下立枯丝核菌菌落直径、菌核形成时间和数量Table 2 Colony diameters ofR.solaniand time and amount of sclerotia formation under different light conditions
光照有利于菌核的形成,在全光照条件下60 h即可形成菌核,菌核的数量显著多于全黑暗和12 h光暗交替条件,培养10 d,全光照培养的菌核数量为493个/皿,光暗交替菌核数量为241个/皿,全黑暗菌核数量为120个/皿,各处理间差异极显著(表2)。
2.6 病菌菌丝及菌核的致死温度
表3结果表明,病菌菌丝经40~48℃处理10 min后,在28℃下培养36 h,在PDA培养基上均能生长,而49℃处理后培养,菌丝完全没有生长,故病菌菌丝致死温度为49℃。
表4结果表明,菌核经40~50℃处理10 min后,在28℃下培养36 h均能萌发形成菌落,经大于51℃的温度处理,菌核均不萌发,故菌核的致死温度为51℃。
3 结论与讨论
本试验结果表明,马铃薯立枯丝核菌菌丝在10~35℃范围内能够生长,但是在低温下菌丝生长缓慢,5℃停止生长,其生长适宜温度为25~30℃;在p H为3~12环境中菌丝均能生长,以p H 7最适生长;在供试的碳氮源中,可溶性淀粉、酵母粉有利于病菌的生长;黑暗有利于菌丝的生长,光照有利于菌核的形成;菌丝的致死温度为49℃,菌核的致死温度为51℃。
李乾坤对甘肃省马铃薯种植区分离的立枯丝核菌的研究表明其菌丝生长温度最低为4℃,最适为23℃,生长上限温度在32~33℃之间,34℃时停止生长[2]。刘宝玉研究表明立枯丝核菌(分离于内蒙古马铃薯产区)菌丝生长适宜温度为24~26℃,最低生长温度为4℃,最高生长温度为34℃;最高生长p H为11[11]。而本试验立枯丝核菌分离于黑龙江九三管理局马铃薯种植田,生物学特性研究表明,该菌5℃即不能生长,生长最适温度为25~30℃,35℃时菌丝生长良好;菌丝在p H为12条件下,也能生长。此结果与前人的研究报道有差异,这可能是分离的菌属于不同的融合群,生物学特性有一定差异。立枯丝核菌的多个融合群可以侵染马铃薯[7-8],因此,应对黑龙江省马铃薯立枯丝核病菌融合群的种类、分布,病菌生理生化、病理遗传等方面进一步研究。

表3 不同高温处理立枯丝核菌后菌落直径Table 3 Colony diameters ofR.solaniat different temperatures

表4 不同高温处理立枯丝核菌菌核后菌落直径Table 4 Colony diameters ofR.solaniat different temperatures
该菌p H适应范围广,但研究表明酸性条件比碱性条件有利于其生长,应属于偏弱酸性菌;可溶性淀粉、蔗糖有利于病菌的生长。因此,在生产上,种植马铃薯时应注意土壤酸碱度以及种植的前茬作物,以减少病害的发生。
[1] 陈万利.马铃薯黑痣病的研究进展[J].中国马铃薯,2012,26(1):49 51.
[2] 李乾坤,孙顺娣,李敏权.马铃薯立枯丝核菌病的研究[J].马铃薯杂志,1988,2(2):79 84.
[3] Caning D E,Baird R E,Gitaitis R D,et al.Characterization of AG-13,a newly reported anastomosis group ofRhizoctonia solani[J].Phytopathology,2002,92(8):893 899.
[4] 李华荣.丝核菌的菌丝融合群及其遗传多样性研究的新进展[J].菌物系统,1999(1):100 107.
[5] 曹春梅,李文刚,张建平,等.马铃薯黑痣病的研究现状[J].中国马铃薯,2009,23(3):171 173.
[6] Scholte K,Lootsma M.Effects of farmyard manure and green manure crops on populations of mycophagous soil fauna andRhizoctoniastem canker of potato[J].Pedobiologia,1996,39(1):15 22.
[7] Virtgen C G,Olalde-Portugal V,Carling D E.Anastomosis groups ofRhizoctonia solanion potato in central Mexico and potential for biological and chemical control[J].American Journal of Potato Research,2000,77(4):219 224.
[8] 王宇,杨志辉,秦宇轩,等.马铃薯黑痣病菌AG2-1和AG3融合群分析[J].农业生物技术学报,2013,21(2):230 237.
[9] 李彩霞.不同杀菌剂防治马铃薯黑痣病田间药效试验[J].农业科技,2012(5):68 69,144.
[10]刘宝玉,蒙美莲,胡俊,等.5种杀菌剂对马铃薯黑痣病的病菌毒力及田间防效[J].中国马铃薯,2010,24(5):306 310.
[11]刘宝玉,胡俊,蒙美莲,等.马铃薯黑痣病病原菌分子鉴定及其生物学特性[J].植物保护学报,2011,28(4):379 380.
Biological characteristics ofRhizoctonia solaniisolated from potato
Tai Lianmei, Zhao Qiaolan, Jin Xuehui, Jin Guanghui
(College of Agronomy,Heilongjiang Bayi Agricultural University,Daqing163319,China)
Biological characteristics ofRhizoctonia solaniwere studied by growth rate method.The results showed that the mycelium could grow under the temperature from 10℃to 35℃;the optimal temperature was 25-30℃,and the fastest growth of mycelia was at 30℃.The culture p H was 3-12,and the optimal p H was 5-7.Darkness could promote the growth of mycelia.Illumination could promote the formation of sclerotia.Various carbon and nitrogen sources significantly influenced the growth of mycelia.Soluble starch and yeast powder had significant promoting effects on the growth of mycelia.The lethal temperatures for mycelia and sclerotia were 49℃and 51℃,respectively.The environmental conditions forR.solaniisolated from potato in Heilongjiang were ascertained,which provides a theoretical basis for control of the disease.
potato;Rhizoctonia solani; biological characteristics
S 435.32
A
10.3969/j.issn.0529 1542.2015.01.017
2014 02 15
2014 05 10
黑龙江省农垦总局攻关项目(HNKXIV-06 03B)
联系方式E-mail:tailianmei@sina.com
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05
中国土壤与肥料(2021年5期)2021-12-02
西北农业学报(2021年5期)2021-06-28
中国食用菌(2021年1期)2021-03-12
昆钢科技(2021年6期)2021-03-09
天津科技(2020年4期)2020-05-09
中国食用菌(2019年9期)2019-10-21
中国海洋大学学报(自然科学版)(2018年11期)2018-10-15
分析化学(2014年10期)2014-10-24
作物研究(2013年2期)2013-09-19
