
重组病毒Ac MNPV-BmK IT侵染Sf9细胞后凋亡相关基因的表达分析
2015-02-14 06:14:48付月君林桃桃梁爱华
植物保护 2015年1期
付月君, 林桃桃, 梁爱华
(山西大学生物技术研究所,化学生物学与分子工程教育部重点实验室,太原 030006)
重组病毒Ac MNPV-BmK IT侵染Sf9细胞后凋亡相关基因的表达分析
付月君*, 林桃桃, 梁爱华
(山西大学生物技术研究所,化学生物学与分子工程教育部重点实验室,太原 030006)
将从东亚钳蝎中克隆到的兴奋型昆虫毒素基因(BmK IT)同源重组到苜蓿银纹夜蛾核型多角体病毒(Ac MNPV)基因组中,得到重组病毒Ac MNPV-BmK IT,抗虫试验表明重组杆状病毒的杀虫活性明显优于野生型病毒,但Ac MNPV介导的BmK IT的抗虫分子机制尚未阐明。本试验从草地贪夜蛾Sf9细胞中克隆获得了凋亡相关基因Sfp53,制备了抗体,分析了Ac MNPV-BmK IT对Sfp53表达的影响,结果表明被重组病毒感染的细胞所表达的Sfp53时间与表达量与野生型相比都有所提前和提高,说明重组病毒可加速细胞的凋亡;同时通过半定量PCR分析了Ac MNPV-BmK IT感染Sf9细胞时病毒抗凋亡基因iap2的表达,结果表明重组型病毒抗凋亡基因iap2表达量减少。以上结果在细胞分子水平上解释了Ac MNPV-BmK IT杀虫活性提高的原因。
Ac MNPV-BmK IT; 凋亡;Sfp53;iap2
蝎使用其毒液作为狩猎和自卫的武器,毒液的有效成分是一组由30~80个氨基酸组成的具有神经毒性的多肽[1-2]。蝎神经毒素按其作用对象大致可分为3类:哺乳动物神经毒素(MTx)、昆虫神经毒素和甲壳动物神经毒素(CTx)[3]。兴奋型昆虫神经毒素的注入可以引起虫体快速兴奋性收缩麻痹[4]。本课题组将从东亚钳蝎中克隆到的兴奋型蝎昆虫毒素基因(BmK IT)与苜蓿银纹夜蛾核型多角体病毒(Autographa californicamulticapsid nucleopolyhedrovirus,Ac MNPV)基因组同源重组,得到重组杆状病毒Ac MNPV-BmK IT,BmK IT基因受极早期启动子IE1 promoter调控。接着,本课题组分别用重组杆状病毒和野生型杆状病毒感染棉铃虫幼虫,结果表明,重组杆状病毒的杀虫活性明显优于野生型杆状病毒,与Fan等[5]报道的结果一致。本研究通过检测细胞凋亡相关基因和病毒抑制凋亡相关基因的表达来分析Ac MNPV介导的BmK IT的抗虫分子机制。
细胞凋亡过程是对细胞死亡的调控,它不仅可以消除多余的正常细胞,也是一种抵抗外源基因的表达与病毒入侵的防御机制[6]。p53蛋白家族参与细胞周期调控、肿瘤的抑制、细胞的凋亡,其可分为无脊椎来源和有脊椎来源[7-9]。Ac MNPV的宿主草地贪夜蛾(Spodoptera frugiperda)细胞中分离出来的Sfp53是一种与鳞翅目昆虫同源的凋亡蛋白。Sfp53的过量表达会诱导细胞的凋亡,但其不会像黑腹果蝇(Drosophila melanogaster)p53基因(Dmp53)那样,受到UV或CPT(camptothecin)处理后,其内源性的Sfp53也会增加[10]。在病毒基因组中,有两种抗凋亡基因:p35与凋亡抑制基因iap(inhibitor of apoptosis)[11]。抗凋亡基因iap首先从苹果蠹蛾颗粒体病毒CpGV(Cydia pomonella granulosis virus)和黄杉毒蛾核型多角体病毒Op MNPV(Orgyia pseudotsugata multiple nucleopolyhedrovirus)中被分离鉴定出来,它所编码的蛋白可以抑制由p35缺失型重组病毒Ac MNPV所诱导的Sf21的凋亡[12-13]。
本试验从草地贪夜蛾Sf9细胞中克隆获得了凋亡蛋白Sfp53基因,构建了pRSETc-Sfp53表达载体,利用Western blot分析了Ac MNPV-BmK IT对细胞凋亡蛋白Sfp53的影响,同时通过半定量PCR分析了Ac MNPV-BmK IT感染Sf9细胞时病毒抗凋亡基因iap2的表达情况,从而为重组病毒Ac MNPV-BmK IT生物杀虫剂的研发及明确其作用机制提供试验依据。
1 材料与方法
1.1 细胞、菌种、质粒、病毒
草地贪夜蛾细胞(Sf9)、大肠杆菌(Escherichia coli)XL10及野生型苜蓿银纹夜蛾核型多角体病毒Ac MNPV由本实验室保存;质粒p RSETc-BmK CT和Ac MNPV-BmK IT由本实验室构建;p MD-18T购自TaKaRa公司。
1.2 主要工具酶及生化试剂
昆虫细胞培养基TNM-FH(Sigma公司);小牛血清(杭州四季青生物工程材料有限公司);RNAisoTMPlus(TaKaRa公司);反转录试剂盒(RevertAid First Strand cDNA Synthesis Kit)、DNA限制性内切酶(Thermo公司);PCR扩增所用EasyTaq酶、T4DNA连接酶和核酸分子量标准(TransGen公司);辣根过氧化物酶标记羊抗兔IgG(Invitrogen公司)。
1.3 昆虫Sf9细胞培养
将50.6 g TNM-FH粉末悬于1 L去离子水中,同时加0.35 g碳酸氢钠,搅拌溶解,调节p H至6.2~6.4,使用灭菌的0.22μm滤膜滤器过滤除菌,配以50μg/mL庆大霉素备用。取出液氮中冻存的Sf9细胞,迅速置于27℃水浴中,待完全融化后1 000 r/min离心5 min,弃去上清液。细胞沉淀用含胎牛血清及抗生素的培养基悬浮,转移至培养面积为25 cm2的细胞培养瓶中,培养基加至5 mL,于27℃细胞培养箱培养。
1.4 试验方法
1.4.1 Sf9细胞总RNA的提取
将Sf9细胞接种于6孔板中,待长满一层细胞时加入1 mL 1×PBS清洗细胞、再加入1 mL RNAisoTMPlus裂解细胞,用移液枪反复吹打细胞使其脱落,将细胞裂解液转移至离心管中反复吹吸,直至裂解液中无明显沉淀后室温静置5 min。向离心管中加入200μL氯仿,用力振荡,待乳化溶液呈乳白状(无分相现象)后室温静置5 min,12 000 g,4℃离心15 min。离心后匀浆液分为3层:无色的上清液、中间的白色蛋白层及下层有机相,吸取上清液转移至另一新的离心管中,向上清中加入等体积的异丙醇,上下颠倒离心管,充分混匀后,在室温下静置10 min,12 000 g,4℃离心10 min,试管底部会出现白色沉淀,弃去上清液,沿离心管壁加入现配的75%冷乙醇l mL,轻轻上下颠倒洗涤离心管管壁,12 000 g,4℃离心5 min后小心弃去乙醇。室温干燥沉淀至没有乙醇残留,加入适量的RNase-free水溶解沉淀,即得Sf9细胞总RNA。
1.4.2 cDNA第一链的合成
采用Revert Aid First Strand cDNA Synthesis Kit,以Oligo d T为引物,将总RNA中的mRNA反转录为cDNA,反应条件为:45℃1 h;70℃10 min;终止反应。
1.4.3Sfp53基因的克隆
根据GenBank数据库中检索到的Spodoptera frugiperda p53 cDNA序列(GenBank accession No.HM773026.1),设计用于扩增Sfp53全长的引物(表1),酶切位点分别为XhoI和NheI(表1中下划线标出)。将cDNA模板稀释10倍,进行PCR扩增。反应条件为:94℃5 min;94℃30 s;53℃30 s;72℃90 s循环30次;72℃10 min。将PCR扩增获得的Sfp53基因构建于克隆载体p MD18-T中,得到p MD18-T-Sfp53,经测序确定其序列正确。

表1 PCR扩增Sfp53和iap2基因所用引物及相关参数Table 1 The primer sequences and relative parameters for PCR amplification ofSfp53 andiap2 genes
1.4.4 重组表达质粒pRSETc-Sfp53的构建
将p MD18-T-Sfp53用NheI和XhoI双酶切后进行1.5%琼脂糖凝胶电泳,回收并纯化得到目的基因Sfp53片段。分别用NheI和XhoI酶切表达载体pRSETc-BmK CT,切胶回收载体pRSETc。连接酶切后的表达载体pRSETc与Sfp53片段(图1)。将测序正确的重组表达质粒pRSETc-Sfp53送生工生物工程(上海)股份有限公司制备Sfp53抗体。
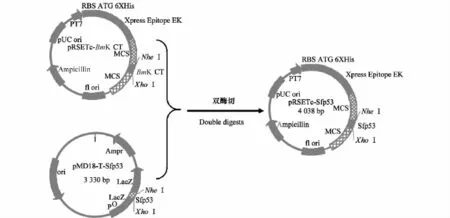
图1 表达质粒pRSETc-Sfp53的构建流程Fig.1 Construction of the expression vector pRSETc-Sfp53
1.4.5 Western blot分析Ac MNPV-BmK IT感染Sf9细胞后Sfp53蛋白的表达
用含血清的昆虫细胞培养基(TNM-FH)将Sf9细胞稀释为5×105个/mL,加2 mL到6孔板中。细胞培养板于27℃培养过夜,次日弃去旧培养基,依次加入无血清培养基(TNM-FH)、2.28×1012vp/ mL的Ac MNPV和Ac MNPV-BmK IT病毒分别感染细胞6、12、21、27、33、39、45 h,共设7个处理,每处理3次重复。提取细胞总蛋白,取5μL用BCA试剂测蛋白浓度,另取总蛋白与Loading buffer按5∶1混匀,金属浴95℃5 min,进行SDS-PAGE电泳。电泳完毕将胶于转膜槽中转印1.5 h。转印完毕,将PVDF膜用PBST清洗后,在封闭液中封闭1 h,期间缓慢摇动。封闭后,将目的蛋白与标准分子量大小进行比较,确定所需目的蛋白位置,剪下所需的膜,将膜用塑料膜封好,留下一小口以便加Sfp53抗体。将抗体稀释2 000倍,从小口中加入,封好,置于4℃孵育过夜。用PBST洗去未结合的一抗,每次10 min,洗涤3次。将PVDF膜在辣根过氧化物酶标记羊抗兔IgG二抗中孵育1 h(二抗稀释比例为1∶6 000),再用PBST洗去未结合的二抗,每次10 min,洗涤3次。在暗室内通过ECL试剂盒曝光显影。
1.4.6 半定量PCR分析Ac MNPV-BmK IT感染Sf9细胞后抗凋亡基因iap2的转录水平
用有血清的昆虫细胞培养基(TNM-FH)将细胞稀释为5×105个/mL,加2 mL到6孔板中,27℃培养箱过夜培养。次日弃去旧培养基,依次加入无血清培养基(TNM-FH),浓度为2.28×1012vp/mL Ac MNPV和Ac MNPV-BmK IT病毒,感染细胞时间为36和60 h,每组3次重复。提取细胞内总RNA,反转录成cDNA。设计用于扩增iap2全长的引物,见表1。将cDNA模板稀释10倍,测定cDNA浓度,保证模板初始量相同条件下进行PCR扩增,反应条件为:94℃5 min;94℃30 s;51℃30 s;72℃1 min循环25次;72℃10 min。等体积PCR产物经1.5%琼脂糖凝胶电泳分析,EB染色。通过BandScan软件对条带灰度进行积分,并进行统计分析。
2 结果与分析
2.1 Sf9细胞总RNA的提取及Sfp53基因的克隆
利用RNAisoTMPlus总RNA提取试剂盒提取Sf9细胞的总RNA,用1%琼脂糖凝胶电泳进行检测,可检测到清晰的28S、18S和5S三条带(图2a),表明所提Sf9的总RNA完整。利用PrimeScriptTMRT Reagent Kit,以Oligo dT为引物合成第一链cDNA。以cDNA为模板,扩增得1 125 bpSfp53序列(图2b),经测序证实扩增目的条带序列正确。

图2 重组表达质粒pRSETc-Sfp53的鉴定Fig.2 Analysis of recombinant plasmid pRSETc-Sfp53
2.2 重组表达质粒pRSETc-Sfp53的鉴定及Sfp53的抗体制备
根据方法1.4.4构建好重组质粒pRSETc-Sfp53,并进行双酶切和PCR鉴定(图2c,d),结果显示质粒构建成功,质粒经测序证实构建正确。将重组质粒交由生工生物工程(上海)股份有限公司进行制备抗体,获得了Sfp53的多克隆抗体。
2.3 Western blot分析Ac MNPV-BmK IT对凋亡
蛋白Sfp53表达量的影响
如图3a所示,重组Ac MNPV-BmK IT感染Sf9细胞12~33 h时,凋亡蛋白Sfp53的表达量增加,39 h以后Sfp53的表达量减少,说明在感染细胞12~33 h时细胞发生凋亡,39 h后细胞凋亡速度减慢,而野生型Ac MNPV感染细胞21 h后,凋亡蛋白Sfp53的表达量才有所增加且增加量不及重组型,感染45 h后Sfp53的表达量小幅度减少,说明野生型Ac MNPV感染Sf9细胞21 h后凋亡主要蛋白Sfp53才开始大量表达,且一直持续到45 h后,凋亡持续时间较长。重组型与野生型病毒处理组相比,前者诱导Sf9细胞的凋亡时间早,持续时间短,加速了细胞凋亡的进程。
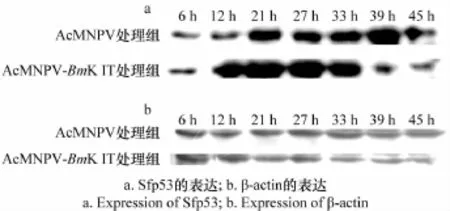
图3 Western blot分析AcMNPV-BmK IT对Sf9细胞凋亡相关蛋白Sfp53表达的影响Fig.3 Effects of Ac MNPV-BmK IT on the expression of apoptosis-related protein Sfp53 by Western blot assay
2.4 半定量PCR分析Ac MNPV-BmK IT感染Sf9细胞后抗凋亡基因iap2的转录水平分析
如图4a所示,与野生型AcMNPV处理组相比,重组AcMNPV-BmK IT感染Sf9细胞后,病毒的抗凋亡基因iap2表达急剧减少,病毒的抗凋亡能力减弱,表明Sf9细胞凋亡被抑制程度减弱。经t检验,同一时间作用下各处理组之间P值小于0.01,差异极显著,说明重组病毒抑制病毒的抗凋亡能力强,加速了宿主细胞的凋亡。

图4 半定量PCR分析AcMNPV-BmK IT感染Sf9细胞对抗凋亡基因iap2转录水平的影响Fig.4 Effects of AcMNPV-BmK IT on the transcription level of the inhibitor of apoptosis geneiap2 by semi-quantitative PCR assay
3 结论与讨论
病毒AcMNPV的复制可以引起Sf9细胞DNA的损伤,同时增加Sfp53的积累;抑制DNA的损伤可以阻止Sfp53的积累,但通过RNA干扰试验沉默Sfp53基因的表达并不影响AcMNPV的复制[14]。本试验表明,AcMNPV-BmK IT可以提高细胞中Sfp53的积累,推测是由于BmK IT通过调控Sf9细胞膜的钠通道,进而影响细胞内环境,从而增强了病毒的复制能力,提高了Sfp53的表达量,表明BmK IT不是从基因水平上影响细胞凋亡的调控,而是在细胞内环境上调控Sfp53蛋白的表达或积累,从而引起细胞凋亡,加强杀虫能力的。
有结果显示Ac MNPV感染Sf9细胞时可以引起凋亡抑制蛋白IAP表达量的降低,IAP下调也可以由病毒DNA的复制所触发,其原因部分是由于某些蛋白酶的介导[15]。与野生型病毒处理组相比,Ac MNPV-BmK IT感染细胞时,iap2的表达得到抑制,推断BmK IT通过调控细胞内环境从而影响了病毒DNA的复制或凋亡,抑制蛋白IAP表达下调酶表达。本试验是从转录水平检测iap2的表达,以后的工作还可开展IAP蛋白表达和IAP表达下调相关酶的检测。有报道发现杆状病毒的IAPs可以阻断Sf-caspase-1的激活[16],本试验结果显示重组病毒的iap2基因的表达大幅下调,这是由BmK IT引起的,下一步工作可分析这种下调对加速细胞凋亡的影响。
本文分析了重组病毒Ac MNPV-BmK IT侵染Sf9细胞后细胞内Sfp53和病毒iap2的表达情况,为重组病毒Ac MNPV-BmK IT生物杀虫剂的研发及作用机制研究提供了试验依据。
[1] Moskowitz H,Hermann R,Jones A D,et al.A depressant insect-selective toxin analog from the venom of the scorpionLeiurus quinquestriatus hebraeu-purification and structure/ function characterization[J].European Journal of Biochemistry,1998,254(1):44 49.
[2] Cestele S,Catterall W A.Molecular mechanisms of neurotoxin action on voltage-gated sodium channels[J].Biochimie,2000,82(9/10):883 892.
[3] Zilberberg N,Gordon D,Elhatem P,et al.Functional expression and genetic alteration of an alpha scorpion neurotoxin[J]. Biochemistry,1996,35(31):10215 10222.
[4] Gurevitz M,Froy O,Zilberberg N,et al.Sodium channel modifiers from scorpion venom:structure-activity relationship,mode of action and application[J].Toxicon,1998,36(11):1671 1682.
[5] Fan X J,Zheng B,Fu Y J,et al.Baculovirus-mediated expression of a Chinese scorpion neurotoxin improves insecticidal efficacy[J].Chinese Science Bulletin,2008,53(12):1855 1860.
[6] Ikeda M,Yanagimoto K,Kobayashi M,et al.Identification and functional analysis ofHyphantria cunea nucleopolyhedrovirus iapgenes[J].Virology,2004,321(2):359 371.
[7] Lane D P,Cheok C F,Brown C J,et al.TheMdm2 andp53 genes are conserved in the Arachnids[J].Cell Cycle,2010,9(4):748 754.
[8] Lu W J,Amatruda J F,Abrams J M.p53 ancestry:gazing through an evolutionary lens[J].Nature Reviews Cancer,2009,9(10):758 762.
[9] Mendoza L,Orozco E,Rodriguez M A,et al.Ehp53,anEntamoeba histolyticaprotein,ancestor of the mammalian tumour suppressor p53[J].Microbiology,2003,149(4):885 893.
[10]Huang N,Clem R J,Rohrmann G F.Characterization of cDNAs encoding p53 ofBombyx moriandSpodoptera frugiperda[J].Insect Biochemistry and Molecular Biology,2011,41(8):613 619.
[11]Clem R J.Baculoviruses and apoptosis:the good,the bad,and the ugly[J].Cell Death and Differentiation,2001,8(2):137 143.
[12]Birnbaum M J,Clem R J,Miller L K.An apoptosis-inhibiting gene from a nuclear polyhedrosis virus encoding a polypeptide with Cys/His sequence motifs[J].Journal of Virology,1994,68(4):2521 2528.
[13]Crook N E,Clem R J,Miller L K.An apoptosis-inhibiting baculovirus gene with a zinc finger-like motif[J].Journal of Virology,1993,67(4):2168 2174.
[14]Huang N,Wu W,Yang K,et al.Baculovirus infection induces a DNA damage response that is required for efficient viral replication[J].Journal of Virology,2011,85(23):12547 12556.
[15]Vandergaast R,Schultz K L,Cerio R J,et al.Active depletion of host cell inhibitor-of-apoptosis IAP triggers apoptosis upon baculovirus DNA replication[J].Journal of Virology,2011,85(16):8348 8358.
[16]Seshagiri S,Miller L K.Baculovirus inhibitors of apoptosis(IAPs)block activation of Sf-caspase-1[J].Proceedings of the National Academy of Sciences U S A,1997,94(25):13606 13611.
Expression analysis of apoptosis-related genes in Sf9 cells infected by recombinant AcMNPV-BmK IT
Fu Yuejun, Lin Taotao, Liang Aihua
(Institute of Biotechnology,Key Laboratory of Chemical Biology and Molecular Engineering
of Ministry of Education,Shanxi University,Taiyuan030006,China)
We engineered an insect toxin gene fromButhus martensiiKarsch(BmK IT gene)into the genome of Ac MNPV(Autographa californicamulticapsid nucleopolyhedrovirus)in our previous study.The bioassay data indicated that the recombinant baculovirus Ac MNPV-BmK IT significantly enhanced the insecticidal activity.However,nothing is known about the apoptosis mechanism of Ac MNPV-BmK IT.In this study,Western blot and semi-quantitative PCR were used to analyze the effects of Ac MNPV-BmK IT on the expression of apoptosis-related genes,Sfp53 andiap2.The results showed that Ac MNPV-BmK IT increased the expression ofSfp53 and inhibited the expression ofiap2,which explained the enhanced insecticidal activity of Ac MNPV-BmK IT in cellular and molecular levels.
Ac MNPV-BmK IT; apoptosis;Sfp53;iap2
Q 781
A
10.3969/j.issn.0529 1542.2015.01.004
2014 07 17
2014 09 01
国家自然科学基金(31272100);山西省自然科学基金(2014011038-1);山西省优秀青年学术带头人支持计划
*通信作者 Tel:0351 7011499,E-mail:yjfu@sxu.edu.cn
猜你喜欢
山东畜牧兽医(2022年10期)2022-12-06 03:52:34
环球时报(2022-09-20)2022-09-20 15:18:57
当代水产(2021年7期)2021-11-04 08:17:54
今日农业(2020年24期)2020-12-15 16:16:00
食品科学(2018年10期)2018-05-23 01:27:28
中国兽医杂志(2016年5期)2016-06-27 01:44:58
兽医导刊(2016年12期)2016-05-17 03:51:50
广东海洋大学学报(2015年4期)2016-01-13 08:39:33
西南医科大学学报(2015年1期)2015-08-22 13:01:46
中国当代医药(2015年9期)2015-03-01 02:01:59
