
奖惩对行为抑制及程序阶段中自主生理反应的影响*
2015-01-24 09:11白学军
心理学报 2015年1期
谷 莉 白学军 王 芹
1 引言
抑制功能是指通过阻止无关信息或干扰信息进入工作记忆中, 而保障个体进行正常思维和行为活动的一种认知加工过程(Nigg, 2000)。行为抑制作为抑制功能的一个重要成分, 与情绪活动之间的交互作用十分复杂(Kalanthroff, Cohen & Henik, 2013;Banich et al., 2009; Guitart-Masip et al., 2012; Rowe,Hirsh, & Anderson, 2007; Yuan, Yang, Meng, Yu, &Li, 2008; Chan, Raine, & Lee, 2010)。有研究显示,被试在正性情绪条件下难以控制优势反应, 说明正性情绪导致行为抑制能力的减弱(朱昭红, 白学军,2009; Chiu, Holmes, & Pizzagalli, 2008; Hare,Tottenham, Davidson, Glover, & Casey, 2005)。但也有研究发现正性情绪可对行为抑制能力产生促进效应, 如 Van der Stigchel, Imants和 Ridderinkhof(2011)采用反向眼跳任务探讨情绪性质与行为抑制能力的关系, 结果显示正性情绪比中性情绪更能降低抑制失败率。相反负性情绪则减弱个体的行为抑制能力(Kalanthroff et al., 2013; 辛勇, 李红, 袁加锦, 2010)。以非正常人群为被试的研究结果也提示行为控制障碍的产生与负性情绪相关。如强迫症个体表现出在惩罚条件下对优势反应的抑制失败率增加, 这种现象和强迫症个体自我报告的症状严重程度成正相关(Morein-Zamir et al., 2013)。而情绪注意偏向的改变则能显著减少强迫行为(Najmi &Amir, 2010)。上述研究结果表明目前有关情绪对行为抑制影响的研究结论并不一致。
Gray的强化敏感性理论(reinforcement sensitivity theory, RST)为情绪与行为抑制的关系提供了生物学解释, Gray认为人脑中存在两个独立作用于行为的情绪系统, 即行为趋近系统(behavior approach system, BAS)和行为抑制系统(behavior inhibition system, BIS)。奖励条件引发高兴、兴奋等正性情绪, 易激活BAS系统, 产生趋近行为; 惩罚条件引发焦虑、恐惧等负性情绪, 易激活BIS系统, 产生回避行为。由于这两个概念性神经系统是Gray以动物实验研究的数据结果概括出来, 在解释特定现象时存在局限性。为此Gray对RST理论进行了修订, BAS系统被解释为对所有欲求性刺激敏感, BIS系统不再对条件的厌恶刺激敏感, 而是负责处理目标冲突。尽管修订后的 RST更适用于解释人类的个体差异, 但仍存在 RST观点无法合理解释研究结果的现象, 因此 RST被认为还不是成熟的理论体系, 需要进一步的理论澄清和实证(Corr, 2001; 郭少聃, 何金莲, 张利燕, 2009), 如一项采用Go/No go任务范式探讨个体冲动性的研究结果显示, 高焦虑者的冲动性高于低焦虑者, 这与RST理论的解释相异, 即焦虑性高的个体应受BIS系统影响表现出更多的抑制行为(Zinbarg &Revelle, 1989)。Corr (2001)在一篇综述中提出, 许多验证 RST理论的研究结果之所以出现预测失败是因为在解释结果时多是用BAS和BIS两系统的独立子系统效应(两个系统是独立的、并可分离的发挥其效应)去解释, 然而独立子系统效应只有在实验条件中不包含混合的奖惩刺激, 也不需要奖惩刺激频繁快速转换, 且个体应对奖励或惩罚刺激极为敏感的情况下才可能发生。目前奖惩相关研究多把奖惩刺激以线索形式呈现, 个体需要根据奖惩线索改变其注意资源的分配, 从而对行为进行快速转换。转换过程中, 奖励与惩罚相互影响、彼此作用,根据奖惩刺激类型或频率等实验条件的不同就会有不一致的实验结果。Corr (2001)认为即便一种刺激也可同时作用于两系统, BAS被激活, BIS就被抑制, 而 BIS被激活, BAS就被抑制, 即联合子系统效应。此外, 一些以生理指标来考察RST理论的研究结果也不尽一致(Bartussek, Diedrich, Naumann,& Collet, 1993; DePascalis, Fiore & Sparita, 1996)。郭少聃等人(2009)认为生理指标更容易接受刺激类型、实验环境及被试对奖惩期待等种种因素的干扰,未来利用实证研究进行理论澄清时, 应选取合适的生理指标, 有必要对强化参数进行精细的操作定义,加强实验控制, 并指出生理指标考察 RST理论的研究结果不一致现象, 是因为心理生理学和神经生理学并非同一个概念, 因此当以具体的神经生理指标去测量心理生理学上概念性神经系统的功能时需要经过一个转换, 这个过程较为复杂。
情绪研究的生理指标中, 与脑电生理及脑功能成像相比, 自主神经的测量所需要的“转换过程”较少, 是生理心理学探讨行为机制较为直接的研究方法。Fowles (2000)在实证研究基础上提出有关奖惩引发情绪与自主神经关系的生理心理学假设, 即奖励条件激活BAS系统, 表现为心率活动的增加; 惩罚条件激活 BIS系统, 则表现为皮肤电活动的增加。但同样有实证研究不支持假设, 如一项探讨ADHD儿童和正常儿童在奖惩线索下自主唤醒特点异同的研究结果发现, 加入惩罚线索后无论是ADHD还是正常儿童的心率都呈现增快现象(Crone,Jennings, & van der Molen, 2003)。这表明BAS系统和BIS系统的生理心理学指标有待更多的实证研究去探讨。本研究除了采集心率和皮肤电指标外, 还同时采集了指脉率和皮温的活动变化。指脉率是通过心脏收缩节律引发的手指血流波浪性的变化, 与心率同样接受自主神经系统两大分支系统的交互作用, 而皮温活动与皮肤毛细血管壁收缩/扩张引起的血流量变化有关, 也可反映自主神经系统的功能活动, 这两个指标都会伴随情绪活动而发生变化。
停止信号任务是评估行为抑制能力的研究范式, 要求被试抑制优势反应而进入非优势反应中。实验程序中包括无停止信号和有停止信号两类任务。因无停止信号任务出现概率较高, 又被称为优势反应; 而有停止信号任务出现概率较低, 又被称为非优势反应(Logan, 1994)。本研究采用停止信号任务探讨奖励与惩罚条件对行为的抑制作用及其自主生理活动的变化。Logan在此研究范式中并没有反馈环节, 因为其研究目的是探究行为抑制功能本身, 无关于奖惩条件。而Corr (2001)指出反馈信号比强化信号更容易被解读成为实际结果, 能充分刺激BAS/BIS系统从而使情绪唤醒更为敏感。本研究借鉴于相关奖惩研究(白学军, 朱昭红, 沈德立,刘楠, 2009; 朱昭红, 白学军, 2009), 在任务结束后呈现给被试正误反馈。
与以往研究有所不同的是, 本研究不仅考察施加奖惩后的自主生理变化, 还包括任务前的准备阶段、任务中的工作阶段及完成任务后的反馈阶段的自主生理变化。已有相关研究多关注于奖惩获得时的自主生理唤醒特点(Crone, Somsen, Zanolie, &Van der Molen, 2006; 白学军等人, 2009; 朱昭红,白学军, 2009), 很少涉及任务准备阶段、工作阶段及正误反馈阶段的自主生理唤醒特点, 这是因为上述研究常把奖惩线索设置为被试内因素, 即在靶刺激呈现之前先呈现一个线索, 来操纵被试可能存在的不同效价的动机偶联性, 因此只考察奖惩获得时的自主生理变化。尽管这种操作式条件反射范式能有效控制个体差异, 但有研究表明情绪刺激出现后需要一定时间才会呈现情绪反应(Dan & Scherer,2008), 这提示短时间内奖惩线索极有可能未引起或未最大限度引起情绪唤醒。此外, 已有研究发现情绪刺激反复呈现时会有重复项反应时下降, 相关脑区激活程度减弱的现象, 表明情绪也具有重复启动效应(Marchewka & Nowicka, 2007; Ishai, Pessoa,Bikle, & Ungerleider, 2004)。一些研究进一步指出不同情绪性质的重复启动效应有显著差异, 如 Hu,Liu和Zhang (2010)的研究结果表明, 尽管中性、正性和负性情绪词都存在重复启动效应, 但正性情绪词的重复启动效应出现最早。Marchewka和Nowicka (2007)的研究结果则发现与中性刺激相比,负性刺激下新项与重复项反应时差异并不显著, 说明负性词并未呈现重复效应。上述研究提示重复性奖励与重复性惩罚引发的情绪体验所带来的行为结果会有所不同。本研究把奖励与惩罚条件作为被试间因素, 既可以避免因频繁更换奖惩线索导致两系统的交互作用, 又可以使奖惩诱发情绪时间较为充分, 而临床教育中更为常见的是累加持续式的奖惩模式, 如对某个体一贯的同一形式的表扬或批评,重复性奖惩的研究结果能为其临床教育提供实证依据和建议。此外, 重复性奖惩所引发的情绪可以作用于完成任务过程中的任意阶段, 因此探讨各阶段生理唤醒特点及程序阶段中的变化既能了解重复性奖惩模式对各程序阶段的影响是否具有差异性, 又能了解哪些自主生理指标的活动更能反应奖励和惩罚间的差别。
按照Gray修订后RST理论和联合子系统效应假说做出以下假设(1)行为变化:由于重复性奖励条件降低被试对奖励的欲求性, BAS系统的激活程度减少, 从而使 BIS系统的抑制功能增强, 导致抑制行为的增加, 而重复性惩罚条件会进一步加剧目标冲突, 从而激活BIS系统, 导致抑制行为的增加; (2)生理变化:奖励条件增加心率变化、指脉率变化和皮温变化, 惩罚条件增加皮肤电变化。
2 方法
2.1 被试
随机选取 18~22岁大学生 45名, 男生 18名,女生27名, 平均年龄为20.39 ± 0.64岁, 所有被试视力正常或矫正视力正常, 无色盲或色弱, 均为右利手。45名被试被随机分成3组, 即奖励组、惩罚组和控制组, 每组15名(其中6名男生、9名女生)。
2.2 实验仪器
实验采用 Super lab系统呈现刺激并记录被试的反应(计时精度为1 ms), 刺激通过DELL 17寸显示器呈现, 显示器显示分辨率为1024×768, 屏幕背景为白色。刺激呈现的屏幕距离被试60 cm。
实验使用16导生理记录系统(BIOPAC MP150)测量被试生理反应, 记录被试在实验期间的心率、指脉率、皮温和皮肤电导反应活动。(1)心率:采用扣式电极以Ⅰ导联方式一端连接于被试, 一端连接于心电描记放大器ECG100G模块上以记录被试的心率变化; (2)指脉率:一端用TSD光敏传感器连接于被试左手拇指指腹处, 一端连接于光电容描记放大器(PPG)以记录被试的指脉率变化; (3)皮肤电导反应:一端用Ag/AgCl电极缠绕被试左手食指和无名指末端指腹处, 一端连接于生理记录系统GSR100C模块上以记录被试的皮肤电导反应的变化; (4)皮温:一端用TSD102热敏电阻传感器连接于被试左手中指指腹处, 一端连接于皮肤温度放大器(SKT100C), 温度单位采用(°F)。本研究将各生理指标的变化定义为不同程序阶段中呈现的生理指标平均值减去各自的基线值。
2.3 实验材料
采用停止信号任务检测被试行为抑制能力。靶刺激为白色云彩下的“雨天”或“雪天”图案, 停止信号为“太阳”, 出现在云彩右上方。出现“雪天”按左键, 出现“雨天”按右键, 出现“太阳”不按键。反馈采用表情图, 答对呈现笑脸, 答错呈现哭脸。奖励与惩罚条件采用金币图片作为刺激物。
2.4 实验程序
实验采用个别施测, 具体实验程序如下:
第一, 实验前准备:上午8点至11点间, 将被试带入室温22度实验室内静坐5 min。
第二, 给被试连接上BIOPAC MP150生理电导仪以记录被试的心率活动、指脉率活动、皮温活动和皮肤电导反应, 为测得基线值, 要求被试保持平静和放松, 持续采集上述生理指标3 min。
第三, 给予被试指导语, 告知被试接下来要进行一个有关注意力的测试。奖励组和惩罚组被试则被告知所获金币数量和完成速度将决定最后所获奖品的等级。两种实验任务需要被试进行按键反应,一种任务要求被试对靶刺激云彩下方的雪天或雨天, 进行快速而正确的左右按键反应, 即无停止信号反应。另一种任务要求被试在看到太阳之后(在靶刺激出现 500 ms后呈现, 且出现在靶刺激“雨天”和“雪天”的几率完全相同)停止按键反应, 称为停止信号反应。实验进程中两种任务随机性呈现。两种实验任务合计70个, 其中无停止信号任务 50个(占任务总数 71.4%), “雨天”和“雪天”靶刺激各25个, 有停止信号20个(占任务总数28.6%)。每个实验任务设置4个程序阶段, 分别是准备阶段、工作阶段、反馈阶段和奖惩阶段。准备阶段屏幕呈现“请准备”; 工作阶段完成停止信号或无停止信号任务; 反馈阶段指在工作阶段后, 正确反应呈现笑脸表情图, 错误反应呈现哭脸表情图; 奖惩阶段中不同组根据反应正误呈现情绪激发图片, 奖励组答对一题屏幕上显示增加一枚金币(最多可获70枚), 答错不减少金币, 惩罚组答错一题屏幕上减少一枚金币(初始显示70枚), 答对不增加金币, 控制组呈现空屏, 在实验程序时间上与奖惩组一致。实验中要求被试快速而正确的进行按键反应。实验任务流程见图1。

图1 实验任务流程图
第四, 进入实验任务的练习阶段, 每个被试都接受练习直至完全熟悉实验要求。
第五, 正式开始实验至实验结束。
2.5 数据采集与分析
停止信号任务存在无停止信号到停止信号的时长(stop-signal delay, SSD)可变与不变的两种形式。一些研究显示这两种形式的停止信号任务所评估的行为抑制效果类似(Sinopoli, Schachar, &Dennis, 2011; Schachar et al., 2007)。本研究中, 由于停止信号在靶刺激出现500ms后呈现(SSD不变),被试在完成无停止信号任务时的反应时中包含了延迟反应时, 因此可把无停止信号的反应时作为评估行为抑制的间接指标。正如Eagle和Baunez(2010)所述, 由于 SSD不变的停止信号反应时没有结束终端而无法直接测量, 所以把无停止信号的反应时和有停止信号的抑制成功(停止按键的正确反应)率作为测量行为抑制的客观指标。鉴于抑制失败率和抑制成功率在统计效果上完全一致(白学军等人,2009), 本研究采用无停止信号的反应时和有停止信号的抑制失败率作为测量行为抑制的行为指标,采集各程序阶段中的心率变化、指脉率变化、皮肤电导反应变化和皮温变化作为测量自主生理反应的活动指标。所有采集的生理数据用AcqKnowledge 4.1软件进行离线分析。
为检验奖惩条件的重复效应, 将实验进程分为三阶段, 其中实验初始 2分钟作为“阶段一” (被试在初始阶段会存在对测试的熟悉过程), 其后实验时间进程平均分为前后两阶段“阶段二”和“阶段三”,两阶段中的无停止信号任务和停止信号任务数量相同。
生理数据采集过程中, 因身体移动及个体差异等因素导致生理数据异常的被试视为无效被试。去除因实验期间频繁移动导致所有生理数据异常的无效被试1人, 去除皮肤电数据中的无效被试3人,皮温数据中的无效被试1人。考虑到快速按键反应的行为数据采集较少受身体移动的影响, 所有 45名被试的行为数据均进入统计分析。
3 结果
3.1 行为指标变化
3.1.1 无停止信号任务的成绩
对奖励组、惩罚组和控制组在无停止信号任务中的反应时进行单因素方差分析, 结果显示, 各组反应时差异显著, F(2,44) = 3.76, p < 0.05, η= 0.2。进一步分析表明, 奖励组(p < 0.05)和惩罚组(p < 0.05)的反应时均显著高于控制组, 奖励组和惩罚组之间的反应时差异不显著。见图 2(a)。此外, 各组在无停止信号任务中的无按键反应百分比为:奖励组10.57 ± 12.95(%); 惩罚组 9.43 ± 10.87(%); 控制组10.29 ± 13.98(%), 单因素方差分析结果显示, 组间无显著差异, F(2,44) = 0.03。
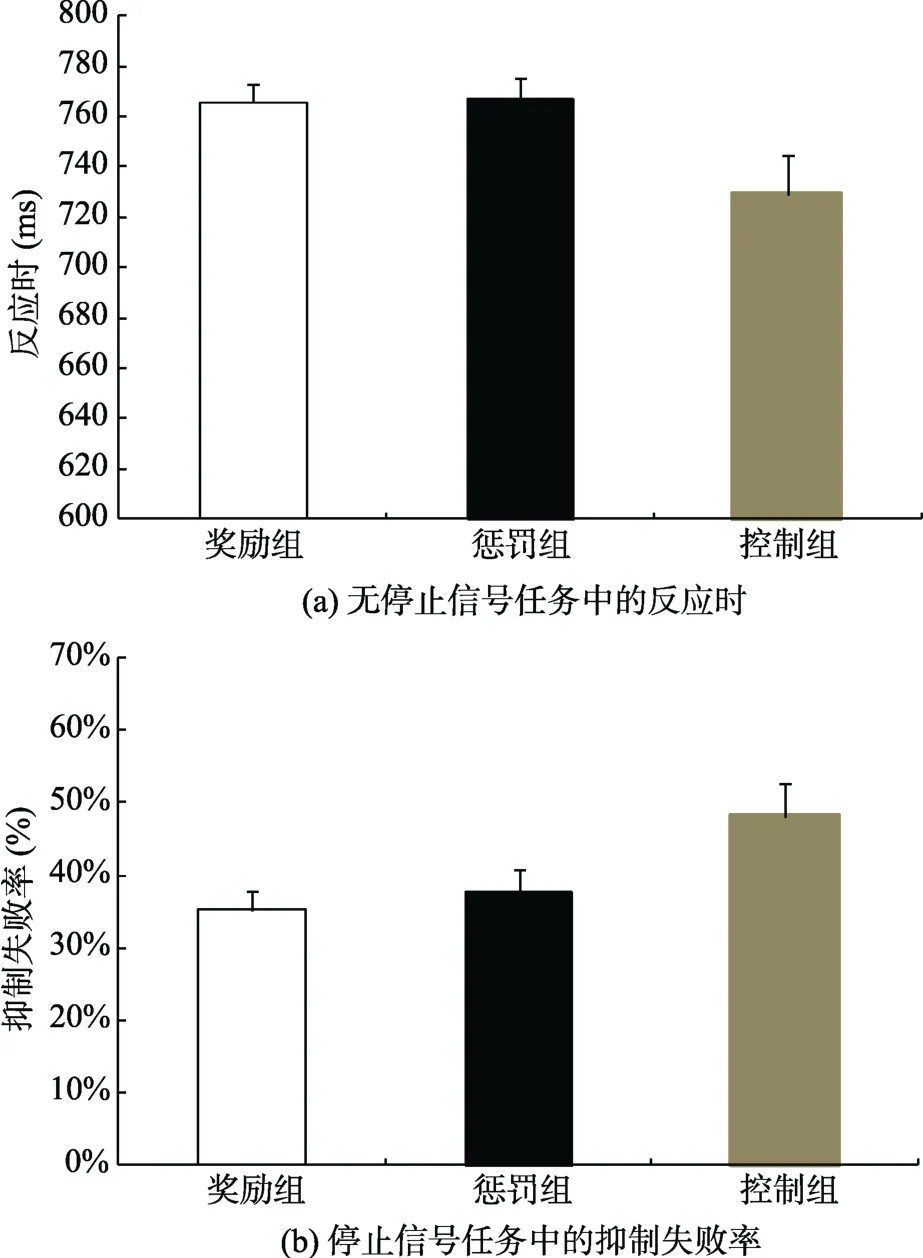
图2 程序阶段中行为指标的变化
3.1.2 有停止信号任务的成绩
对奖励组、惩罚组和控制组在有停止信号任务中的抑制失败率进行单因素方差分析, 结果显示,各组抑制失败率差异显著, F(2,44) = 3.78, p < 0.05,η= 0.2。进一步分析表明, 奖励组(p < 0.05)和惩罚组(p < 0.05)的抑制失败率均显著低于控制组, 奖励组和惩罚组之间的抑制失败率差异不显著。见图2(b)。
3.1.3 反应时与抑制失败率的重复启动效应
对各条件下三阶段中的反应时和抑制失败率的重复效应采用3(条件:奖励、惩罚、控制)×3(阶段一、阶段二、阶段三)的重复方差分析。反应时结果显示, 时间进程主效应显著, F(2,84) = 20.28, p <0.001, η= 0.33, 进一步分析表明阶段一的反应时显著高于阶段二(p < 0.001)和阶段三(p < 0.001)。奖惩条件主效应不显著, F(2,42) = 2.03; 奖惩条件和时间进程交互作用显著, F(4,84) = 2.82, p < 0.05, η=0.12, 简单效应分析结果显示, 阶段二中各组反应时差异显著(p < 0.01), Bonferroni事后检验显示,奖励组的反应时显著高于控制组(p < 0.01)。重复测量方差分析结果显示, 奖励组(p < 0.001)、惩罚组(p <0.05)和控制组(p < 0.05)的反应时时间进程变化显著, Bonferroni检验结果显示, 奖励组阶段一的反应时显著低于阶段二(p < 0.001)和阶段三(p < 0.05),而阶段二的反应时则显著高于阶段三(p < 0.01)。见图 3(a)。

图3 实验时间进程中奖惩条件的重复启动效应
抑制失败率结果显示, 时间进程主效应显著,F(2,84) = 69.05, p < 0.001, η= 0.62, 进一步分析表明阶段一的抑制失败率显著高于阶段二(p < 0.001)和阶段三(p < 0.001)。奖惩条件主效应不显著,F(2,42) = 0.58; 奖惩条件和时间进程交互作用显著,F(4,84) = 5.54, p < 0.01, η= 0.21, 简单效应分析结果显示, 阶段二中各组抑制失败率差异显著(p <0.01), Bonferroni事后检验显示, 奖励组的抑制失败率显著低于惩罚组(p < 0.01)和控制组(p <0.001)。重复测量方差分析结果显示重复测量方差分析结果显示, 奖励组(p < 0.001)、惩罚组(p <0.001)和控制组(p < 0.001)的抑制失败率随时间进程变化显著, Bonferroni检验结果显示, 奖励组阶段一的抑制失败率显著高于阶段二(p < 0.001)和阶段三(p < 0.001), 而阶段二的抑制失败率则显著高于阶段三(p < 0.01); 惩罚组阶段一的抑制失败率显著高于阶段二(p < 0.01)和阶段三(p < 0.001); 控制组阶段一的抑制失败率显著高于阶段二(p < 0.05)和阶段三(p < 0.01), 而阶段二的抑制失败率则显著高于阶段三(p < 0.05)。见图3(b)。
3.1.4 反应时与抑制失败率的相关统计
分别对各组的有停止信号任务的反应时和无停止信号任务的抑制失败率进行皮尔逊(Pearson)相关分析, 结果发现, 惩罚组的反应时与抑制失败率在0.01水平上呈显著负相关(r = −0.76), 控制组的反应时与抑制失败率在0.001水平上也呈显著负相关(r = −0.87), 而奖励组的反应时与抑制失败率则显示不相关(r = −0.27)。
3.2 程序阶段中的生理指标变化
对各组被试在各程序阶段中的生理指标变化,采用3(奖惩条件:奖励组、惩罚组、控制组)×4(程序阶段:准备阶段、工作阶段、反馈阶段、奖惩阶段)的混合设计模式进行分析。
3.2.1 心率变化
心率变化的奖惩条件主效应显著, F(2,41) =5.08, p < 0.05, η= 0.2, 进一步分析显示, 奖励组的心率变化显著高于惩罚组(p < 0.05)和控制组(p <0.01)。程序阶段主效应显著, F(3,123) = 4.48, p <0.01, η= 0.1, 表明不同程序阶段下个体的心率变化差异显著, 进一步分析显示, 反馈阶段的心率变化显著小于工作阶段(p < 0.05)和奖惩阶段(p <0.001), 准备阶段的心率变化显著小于奖惩阶段(p< 0.05)。奖惩条件和程序阶段之间交互作用显著,F(6,123) = 3.8, p < 0.01, η= 0.16, 简单效应分析表明, 各程序阶段下奖励与惩罚条件引起的心率变化差异均显著(准备阶段p < 0.01; 工作阶段p < 0.01;反馈阶段p < 0.05; 奖惩阶段p < 0.05), LSD事后检验表明, 除反馈阶段奖励组和惩罚组之间差异不显著, 其它各阶段的奖励组的心率变化均显著高于惩罚组和控制组。见图4(a)。

图4 程序阶段中生理指标的变化
3.2.2 皮温变化
皮温变化的奖惩条件主效应显著, F(2,40) =3.28, p < 0.05, η= 0.14, 进一步分析显示, 奖励组的皮温变化显著高于控制组(p < 0.05)。程序阶段主效应不显著。奖惩条件和程序阶段之间交互作用显著, F(6,120) = 3.51, p < 0.01, η= 0.15, 简单效应分析表明, 除反馈阶段外, 在准备阶段(p < 0.05)、工作阶段(p < 0.05)和奖惩阶段(p < 0.05)中皮温变化差异均显著, LSD事后检验表明, 各阶段的奖励组的皮温变化均显著高于控制组。此外, 控制组的One-Sample t Test结果显示, 除反馈阶段外, 各程序阶段中的皮温变化相对于基线均显著降低(p <0.05), 而惩罚条件下各程序阶段的皮温变化均未显示出差异性。见图4(b)。
3.2.3 指脉率变化
指脉率变化的奖惩条件主效应不显著, 程序阶段主效应不显著。奖惩条件和程序阶段之间交互作用显著, F(6,123) = 6.31, p < 0.001, η= 0.24, 简单效应分析表明, 除反馈阶段外, 在准备阶段(p <0.05)、工作阶段(p < 0.05)和奖惩阶段(p < 0.05)下奖励与惩罚条件引起的皮温变化差异均显著, LSD事后检验表明, 各阶段的奖励组的皮温变化均显著高于控制组。见图4(c)。
3.2.4 皮肤电变化
皮肤电变化的奖惩条件主效应显著, F(2,38) =3.53, p < 0.05, η= 0.16, 进一步分析显示, 奖励组皮肤电变化显著小于惩罚组(p < 0.05)和控制组(p <0.05)。程序阶段主效应显著, F(3,114) = 15.86, p <0.001, η= 0.29, 表明不同程序阶段中个体的皮肤电变化差异显著, 进一步分析显示, 奖惩阶段显著高于准备阶段(p < 0.01)、工作阶段(p < 0.001)和反馈阶段(p < 0.001); 准备阶段显著高于工作阶段(p< 0.001)和反馈阶段(p < 0.001)。奖惩条件和程序阶段之间交互作用不显著。见图4(d)。
3.3 行为指标与生理指标的相关分析
为验证行为唤醒与生理唤醒之间是否存在相关性, 我们分别对各组的各生理指标在不同程序阶段下与各行为指标(停止信号反应时和抑制失败率)进行皮尔逊相关分析, 结果发现, 各组的各生理指标与行为指标均无相关。
4 讨论
4.1 奖励与惩罚条件均对行为产生抑制作用
本研究行为数据结果显示无论是奖励条件还是惩罚条件, 均可使停止信号任务的抑制失败率降低, 表明从抑制结果上看, 奖励与惩罚在重复性条件下对行为抑制能力均具有促进作用。另一方面,无论是奖励条件还是惩罚条件, 都可使无停止信号任务的反应时增加, 说明从行为抑制的发生过程上看, 奖励条件和惩罚条件均对行为活动产生抑制作用。各组停止信号任务反应时和无停止信号任务抑制失败率的相关分析结果显示, 惩罚组和控制组的反应时与抑制失败率均呈现显著负相关, 而惩罚组与控制组相比, 其抑制失败率显著降低, 说明控制组存在因没有奖惩刺激而快速按键导致失败率增加的可能性, 而惩罚条件使被试为避免接受惩罚通过增加反应时降低抑制失败率。但奖励组的反应时与抑制失败率则呈现不相关, 表明被试在奖励条件下呈现的反应时增加和抑制失败率降低并无关系,每一次任务完成后的正性反馈并没有驱使被试为追求成功率而增加反应时时间。奖惩组的相关差异提示奖励与惩罚条件虽然都显示出对行为活动的抑制作用, 但其发生机制仍具有差异性, 这应与奖惩刺激的重复性有关。如引言部分所述, 正性刺激更容易产生重复启动效应, 相反, 负性刺激则不易产生重复启动效应(Hu et al., 2010; Marchewka &Nowicka, 2007)。本研究关于奖惩条件重复启动效应的统计结果显示, 与反应时逐阶段增加、抑制失败率逐阶段降低的惩罚组和控制组相比, 奖励组在阶段二呈现了反应时增加和抑制失败率减少的显著差异, 而在阶段三则又呈现出回落现象, 结果不仅表明奖励组被试可在阶段时间内提高行为抑制能力, 同时也表明奖励条件下的情绪重复启动效应更大。情绪的重复启动效应差异性可以从“情绪负性偏向”来解释, 即个体对环境中的负性情绪更具有敏感性, 负性信息能获取更快更多的注意资源(Ohman, Lundqvist, & Esteves, 2001; Vaish, Grossmann, & Woodward, 2008)。如果习惯了重复接收负性刺激, 将导致个体警戒下降、减少分配到威胁信号上的注意资源, 负性刺激一旦发生变化时, 个体存在无法及时防御而受到伤害的可能性。因此, 奖励条件对行为的抑制作用是因为重复性奖励降低了被试对奖励的欲求性, 而惩罚条件对行为的抑制作用是其情绪性质本身所产生的结果。
我们根据 Gray修订后的理论及联合子系统效应假说尝试对本研究结果进行解释, 修订后的BAS系统被认为对所有欲求刺激敏感, BIS系统则被认为主要负责处理目标冲突, 当呈现引发目标冲突的刺激时, BIS被激活, 从而抑制在冲突中本来占优势的行为。我们认为, 由于重复性奖励条件一定程度上降低被试对奖励的欲求性, 因此降低BAS的激活程度, 从而减弱对BIS的抑制作用, BIS激活程度增加, 导致抑制行为的增加; 重复性惩罚条件由于受重复启动效应较小, 仍对惩罚刺激较为敏感, 使得BIS系统激活程度增加。本研究范式中对优势反应的抑制过程是一种目标冲突刺激, 会一定程度的激活BIS系统, 所以在本实验条件下不仅惩罚条件, 奖励条件也会伴有BIS系统一定程度的激活, 从而在任务过程中表现出抑制行为的增加。研究结果提示, 尽管奖惩条件都可使抑制失败率降低, 但从发生过程上看, 奖惩条件又均会产生行为抑制, 而教育活动中的行为抑制将影响个体的学习主动性和灵活性。因此, 临床教育中应注意对个体的奖惩实施策略, 重复性奖励会使个体降低对奖励的欲求性, 减少奖励的激励效应, 应避免相同形式的重复性奖励; 重复性惩罚则不会降低个体对惩罚的敏感性从而对行为产生抑制作用, 也应在临床教育中避免使用。
4.2 奖励与惩罚条件对自主生理活动的影响及程序阶段中自主生理活动变化的影响
现代情绪理论认为自主神经反应不仅是情绪的主要功能之一, 并且能对特定情绪情境产生具有预测性行为的生理反应。尽管本研究的行为结果显示, 奖励与惩罚条件均能引发行为抑制, 但奖惩条件对自主生理活动的影响则有显著差异。生理指标与行为指标的相关分析结果显示, 不同程序阶段中各组的各生理指标与行为指标均无相关, 说明行为唤醒与生理唤醒并不同步, 可能存在分离现象。一项讨论主观报告、生理和行为之间关系的研究结果同样显示, 生理活动的变化与主观报告及表情行为之间存在较低相关度(Mauss, Levenson, McCarter,Wilhelm, & Gross, 2005)。正如一些研究者所述, 行为唤醒和生理唤醒并非总是同步发生(Petri &Govern, 2004/2005; Quas, Hong, Alkon, & Boyce,2000)。Petri和Govern (2005)等人指出尽管唤醒理论的基本假设是, 我们可以通过监控脑的活动或自主神经系统的变化, 将它们与行为的变化相联系,以此对唤醒进行心理生理测量, 而这些指标之间实际上缺乏实质性相关, 因为行为唤醒可看作是通过有机体的反应来表现, 自主唤醒可以通过身体机能的变化来加以证实, 虽然它们可能会同时发生, 但它们不一定必然同时发生, 事实上它们是相互独立的。
生理指标奖惩效应的统计结果显示, 奖励组被试的心率活动显著高于惩罚组和控制组、皮温活动显著高于控制组; 而奖励组的皮肤电活动则显著低于惩罚组和控制组。这些生理活动的变化不仅呈现出奖惩条件对自主神经活动的影响模式, 也提示不同生理指标的奖惩效应有所不同。心率指标的结果验证了Fowles关于BAS的生理心理学假设, 即心率活动是 BAS激活标志的生理心理学指标。这可以考虑为 BAS系统在对趋近行为的调节过程中,无论是心输出量的增加和还是能量需求上的增加,都表现出与交感神经系统的功能支配更为相关, 因此奖励条件通过激活 BAS系统增强交感神经系统的兴奋性从而促进心率增加。皮温活动的统计结果显示, 奖励组的皮温活动显著高于控制组。我们认为这与被试在奖惩条件下的按键反应有关, 表明皮温活动在奖励条件驱使下更为关注其任务过程中的按键反应, 在交感神经系统的功能调节下手指皮肤的舒血管纤维兴奋性提高, 毛细血管舒张, 指端血流量增加, 从而使皮温升高; 控制组的皮温相对于基线有所降低, 而惩罚组则没有显著差异变化,则说明伴随实验进程控制组被试减少了对手指按键反应的关注从而导致皮温降低, 而重复性惩罚条件持续作用于指温度变化, 使之始终保持在同一水平。指脉率指标的奖惩效应并不显著, 提示指脉率似乎不易接受奖惩条件的影响。王振宏、郭德俊、游旭群和高培霞(2007)利用情绪生理学方法探讨了身体攻击行为学生和普通学生的情绪生理唤醒特点, 厌恶、恐惧、悲伤、愉快诱发刺激既不能引起普通学生指脉率的显著变化, 也不能引起身体攻击行为学生指脉率的显著变化, 说明指脉率与其它生理指标相比对情绪生理唤醒较不敏感。皮肤电活动通过汗腺活动变化从而影响和改变皮肤电阻变化,人的生理心理反应性汗腺分泌主要接受交感神经系统的功能支配。本研究中惩罚组的皮肤电活动并没有增加, 这与Fowels所提出体现BIS系统被激活的生理心理学指标为皮肤电的研究假说并不一致,我们认为这可能与整体实验时间较长未体现出其变化特点相关。相反, 奖励组的皮肤电活动显著降低则验证了躯体标识假设(Somatic marker hypothesis)。这种假设提出皮肤电活动反应的高低是冒险性选择信号的标识, 较低的皮肤电活动提示个体倾向于冒险性行为, 较高的皮肤电活动则提示个体对情绪性刺激所做出的适宜性行为反应(Fishbein et al., 2005)。冒险性行为的趋近特点应体现出对BAS系统的激活, 因此在本研究中不仅是心率活动, 皮肤电活动也可作为BAS系统的生理心理学指标。
生理指标在各程序阶段中的统计结果显示, 心率和皮肤电指标在反馈阶段的变化最少; 皮温和指脉率指标则在反馈阶段中没有显示出奖惩条件下的显著变化, 而其它阶段都呈现出奖惩效价。这提示与其它程序阶段相比, 反馈阶段的生理指标变化最小, 接受重复性奖惩的影响较少。本实验程序中反馈阶段呈现笑脸或哭脸以提示被试任务完成的正确与否, 呈现时间为 1000 ms。一种可能性为反馈阶段的情绪唤醒程度较低, 另一种可能性正如前言所述, 情绪反应需要一定时间, 短时间的反馈阶段不容易呈现情绪引发的生理变化, 这给选取合适程序阶段采集情绪生理指标的相关研究提供一定参考。此外, 皮肤电的程序阶段变化较为显著, 表现为奖惩阶段的皮肤电变化显著高于其它阶段, 这与Sohn, Sokhadze和Watanuki (2001)对皮肤电活动特点的表述较为一致, 即皮肤电活动对情绪唤醒最为敏感。Kreibig, Wilhelm, Roth和Gross (2007)解释为汗腺分泌影响皮肤电导反应, 情绪唤醒特别会引发手部汗液增加的现象, 因此皮肤电活动应是一项对情绪唤醒非常灵敏的生理指标。本研究中由于各程序阶段的情绪唤醒程度有所不同, 实验设置在奖惩阶段的情绪唤醒程度最高, 因此皮肤电活动的变化在奖惩阶段最为显著。此外, 与其它生理指标相比, 各组的皮肤电指标呈现出一致性的程序阶段反应, 各程序阶段中的情绪效价差异几乎没有发生变化, 这说明皮肤电活动的情绪效价较为稳定, 较少接受程序阶段变化的影响。
5 结论
在本实验条件下, 可得出以下结论:奖励与惩罚条件对行为抑制的行为和生理活动的影响是不同步的; 奖励与惩罚条件均对行为有抑制作用; 奖励条件下被试的心率活动增加, 皮肤电活动降低;各生理指标在准备、工作和奖惩阶段中的奖惩差异性显著。
Bai, X. J., Zhu, Z. H., Shen, D. L., & Liu, N. (2009).Autonomic nervous arousal and behavioral response of punishment and reward in extroverts and introverts. Acta Psychologica Sinica, 41(6), 492–500.
[白学军, 朱昭红, 沈德立, 刘楠. (2009). 奖惩线索条件下内外倾个体的自主唤醒和行为反应. 心理学报, 41(6),492–500.]
Bartussek, D., Diedrich, O., Naumann, E., & Collet, W. (1993).Introversion-extraversion and Event-Related Potential (ERP):A test of J.A. Gray's theory. Personality and Individual Differences, 14, 565–574.
Banich, M. T., Mackiewicz, K. L., Depue, B. E., Whitmer, A.J., Miller, G. A., & Heller, W. (2009). Cognitive control mechanisms, emotion and memory: A neural perspective with implications for psychopathology. Neuroscience and Biobehavioral Reviews, 33(5), 613–630.
Chan, S. C., Raine, A., & Lee, T. M. (2010). Attentional bias towards negative affect stimuli and reactive aggression in male batterers. Psychiatry Research, 176(2–3), 246–249.
Chiu, P. H., Holmes, A. J., & Pizzagalli, D. A. (2008).Dissociable recruitment of rostral anterior cingulate and inferior frontal cortex in emotional response inhibition.NeuroImage, 42(2), 988–997.
Crone, E. A., Jennings, J. R., & van der Molen, M. W. (2003).Sensitivity to interference and response contingencies in attention-deficit/hyperactivity disorder. Journal of Child Psychology and Psychiatry, and Allied Disciplines, 44(2),214–226.
Crone, E. A., Somsen, R. J., Zanolie, K., & Van der Molen, M.W. (2006). A heart rate analysis of developmental change in feedback processing and rule shifting from childhood to early adulthood. Journal of Experimental Child Psychology,95(2), 99–116.
Corr, P. J. (2001). Testing problems in J. A. Gray’s personality theory: A commentary on Matthews and Gilliland.Personality and Individual Differences, 30, 333–352.
Dan, G. E., & Scherer, K. R. (2008). Neuronal processes involved in subjective feeling emergence: Oscillatory activity during an emotional monitoring task. Brain Topography,20(4), 224–231.
DePascalis, V., Fiore, A. D., & Sparita, A. (1996). Personality,event-related potential (ERP) and heart rate (HR): An investigation of Gray's theory. Personality and Individual Differences, 20, 733–746.
Eagle, D. M., & Baunez, C. (2010). Is there an inhibitoryresponse-control system in the rat? Evidence from anatomical and pharmacological studies of behavioral inhibition.Neuroscience and Biobehavioral Reviews, 34(1), 50–72.
Fishbein, D., Hyde, C., Eldreth, D., London, E. D., Matochik,J., Ernst, M., Isenberg, N., … Kimes, A. (2005). Cognitive performance and autonomic reactivity in abstinent drug abusers and nonusers. Experimental and Clinical Psychopharmacology, 13(1), 25–40.
Fowles, D. C. (2000). Electrodermal hyporeactivity and antisocial behavior: Does anxiety mediate the relationship?Journal of Affective Disorders, 61(3), 177–189.
Guitart-Masip, M., Huys, Q. J., Fuentemilla, L., Dayan, P.,Duzel, E., & Dolan, R. J. (2012). Go and no-go learning in reward and punishment: Interactions between affect and effect. NeuroImage, 62(1), 154–166.
Guo, S. D., He, J. L., & Zhang, L. Y. (2009). Reinforcement sensitivity theory of personality: A commentary. Advances in Psychological Science, 17(2), 390–395.
[郭少聃, 何金莲, 张利燕. (2009). 强化敏感性人格理论述评. 心理科学进展, 17(2), 390–395.]
Hare, T. A., Tottenham, N., Davidson, M. C., Glover, G. H., &Casey, B. J. (2005). Contributions of amygdala and striatal activity in emotion regulation. Biological Psychiatry, 57(6),624–632.
Hu, Z., Liu, H., & Zhang, J. X. (2010). Effects of material emotional valence on the time course of massive repetition priming. Journal of Psycholinguistic Research, 39(3),199–211.
Ishai, A., Pessoa, L., Bikle, P. C., & Ungerleider, L. G. (2004).Repetition suppression of faces is modulated by emotion.Proceedings of National Academy of Sciences of United States of America of the United States of America, 101(26),9827–9832.
Kalanthroff, E., Cohen, N., & Henik, A. (2013). Stop feeling:Inhibition of emotional interference following stop-signal trials. Frontiers in Human Neuroscience, 7, 78.
Kreibig, S. D., Wilhelm, F. H., Roth, W. T., & Gross, J. J.(2007). Cardiovascular, electrodermal, and respiratory response patterns to fear- and sadness-inducing films. Psychophysiology, 44(5), 787–806.
Logan, G. D. (1994). On the ability to inhibit thought and action: A user’s guide to the stop signal paradigm. In D.Dagenbach & T. H. Carr (Eds.), Inhibitory processes in attention, memory, and language (pp. 189–239). San Diego:Academic Press.
Marchewka, A., & Nowicka, A. (2007). Emotionally negative stimuli are resistant to repetition priming. Acta Neurobiologiae Experimentalis, 67(1), 83–92
Mauss, I. B., Levenson, R. W., McCarter, L., Wilhelm, F. H.,& Gross, J. J. (2005). The tie that binds? Coherence among emotion experience, behavior, and physiology. Emotion,5(2), 175–190.
Morein-Zamir, S., Papmeyer, M., Gillan, C. M., Crockett, M.J., Fineberg, N. A., Sahakian, B. J., …Robbins, T. W.(2013). Punishment promotes response control deficits in obsessive-compulsive disorder: Evidence from a motivational go/no-go task. Psychological Medicine, 43(2), 391–400.
Najmi, S., & Amir, N. (2010). The effect of attention training on a behavioral test of contamination fears in individuals with subclinical obsessive-compulsive symptoms. Journal of Abnormal Psychology, 119(1), 136–142.
Nigg, J. T. (2000). On inhibition/disinhibition in developmental psychopathology: Views from cognitive and personality psychology and a working inhibition taxonomy.Psychological Bulletin, 126(2), 220–246.
Ohman, A., Lundqvist, D., & Esteves, F. (2001). The face in the crowd revisited: A threat advantage with schematic stimuli. Journal of Personality and Social Psychology,80(3), 381–396.
Petri, H. L., & Govern, J. M. (2005). Motivation theory,research, and applications (Y. B. Guo, Trans.). Xi’an:Shanxi Normal University Press. (Original work published 2004)
[Petri, H. L., & Govern, J. M.. (2005). 动机心理学 (郭本禹译). 西安: 陕西师范大学出版社.]
Quas, J. A., Hong, M., Alkon, A., & Boyce, W. T. (2000).Dissociations between psychobiologic reactivity and emotional expression in children. Developmental Psychobiology,37(3), 153–175.
Rowe, G., Hirsh, J. B., & Anderson, A. K. (2007). Positive affect increases the breadth of attentional selection.Proceedings of the National Academy of Sciences of the United States of America, 104(1), 383–388.
Schachar, R., Logan, G. D., Robaey, P., Chen, S., Ickowicz, A.,& Barr, C. (2007). Restraint and cancellation: Multiple inhibition deficits in attention deficit hyperactivity disorder.Journal Abnormal Child Psychology, 35(2), 229–238.
Sinopoli, K. J., Schachar, R., & Dennis, M. (2011). Reward improves cancellation and restraint inhibition across childhood and adolescence. Developmental Psychology,47(5), 1479–1489.Sohn, J. H., Sokhadze, E., & Watanuki, S. (2001). Electrodermal and cardiovascular manifestations of emotions in children. Journal of Physiological Anthropology and Applied Human Science, 20(2), 55–64.
Vaish, A., Grossmann, T., & Woodward, A. (2008). Not all emotions are created equal: The negativity bias in socialemotional development. Psychological Bulletin, 134(3),383–403.
Van der Stigchel, S., Imants, P., & Ridderinkhof, K. R. (2011).Positive affect increases cognitive control in the antisaccade task. Brain and Cognition, 75(2), 177–181.
Wang, Z. H., Guo, D. J., You, X. Q., & Gao, P. X. (2007).Emotional arousal of autonomic nervous activity of students with physical aggressive behavior. Acta Psychologica Sinica,39(2), 277–284.
[王振宏, 郭德俊, 游旭群, 高培霞. (2007). 身体攻击行为学生自主神经活动的情绪唤醒特点. 心理学报, 39(2),277–284.]
Xin, Y., Li, H., & Yuan, J. J. (2010). Negative emotion interferes with behavioral inhibitory control: An ERP study.Acta Psychologica Sinica, 42(3), 334–341.
[辛勇, 李红, 袁加锦. (2010). 负性情绪干扰行为抑制控制:一项事件相关电位研究. 心理学报, 42(3), 334–341.]
Yuan, J. J., Yang, J. M., Meng, X. X., Yu, F. Q., & Li, H.(2008). The valence strength of negative stimuli modulates visual novelty processing: Electrophysiological evidence from an event-related potential study. Neuroscience, 157(3),524–531.
Zhu, Z. H., & Bai, X. J. (2009). The effect of reward and punishment on response disinhibition, heart rate and skin conductance level during the stop signal task. Psychological Science, 32(4), 796–800.
[朱昭红, 白学军. (2009). 奖惩线索对停止信号任务中抑制能力和心率、皮肤电的影响. 心理科学, 32(4), 796–800.]
Zinbarg, R., & Revelle, W. (1989). Personality and conditioning: A test of four models. Journal of Personality and Social Psychology, 57(2), 301–314.
猜你喜欢
数字技术与应用(2022年3期)2022-04-14
口腔医学(2020年6期)2020-07-08
爱你(2019年21期)2019-11-14
青少年科技博览(中学版)(2019年7期)2019-10-11
爱你·健康读本(2019年6期)2019-07-08
文教资料(2019年31期)2019-01-14
长江丛刊(2017年10期)2017-11-24
赤峰学院学报·哲学社会科学版(2016年12期)2017-03-20
校园英语·中旬(2016年8期)2016-07-09
中国实用医药(2015年24期)2015-05-08
