
辽宁省新城疫病毒F基因和H N基因的分子特征与致病性
2014-08-08 06:13赵晓彤吴运谱薛树山曹明慧顾贵波谷志大王宏燕闫海滨于长泳
中国兽医杂志 2014年3期
赵晓彤,吴运谱,薛树山,石 霖,曹明慧,顾贵波,谷志大,魏 澍,何 欣,王宏燕,闫海滨,于长泳,曹 东
(1.辽宁省动物疫病预防控制中心,辽宁 沈阳 110164;2.辽宁省动物医学研究院,辽宁 沈阳 110164;3.中国兽医药品监察所,北京 海淀 100081)
新城疫(ND)是由新城疫病毒(NDV)所引起的禽类一种急性、烈性传染病,其主要侵害鸡和火鸡,发病率和死亡率均非常高,为危害我国养禽业的最严重疾病之一[1],世界动物卫生组织(OIE)规定为A类动物疫病,我国将其列为一类动物疫病,为国家中长期动物疫病防治规划(2012—2020年)所规定的5种优先防治的一类动物疫病之一。
依据基因组长度不同,NDV分为Class I和Class II两大谱系[1],Class I型的基因组长度为15198nt,Class II中I-IV型和V-IX型NDV分别为15186nt和15192nt。NDV虽只有一个血清型,但其毒株众多且毒力差别较大[1-3],给ND的防制带来极大困难。NDV为单股不分节段负链RNA病毒,病毒基因组结构模式为3'-NPP-M-F-HN-L-5',其囊膜表面糖蛋白-融合蛋白(F)介导病毒囊膜与靶细胞膜融合,促使病毒基因组进入胞桨[4],为决定NDV毒力的主要因素之一。血凝素-神经氨酸酶(HN)能够识别靶细胞的唾液酸受体,介导病毒粒子对靶细胞的吸附,促使新生病毒粒子从感染细胞膜表面释放,并促进融合蛋白的融合功能[5]。因此对辽宁NDV分离株的F、HN基因进化规律的研究,可为本省制定新城疫防制对策提供依据。
1 材料与方法
1.1 SPF鸡胚 9-11日龄SPF鸡胚购自辽宁省益康生物股份有限公司。
1.2 主要试剂 NDV标准抗原及标准阳性血清、H5、H7、H9亚型禽流感标准阳性血清,购自哈尔滨维科公司;EDS76血凝抗原和HI试验阳性血清,购自中国兽医药品监察所;NDV核酸荧光RT-PCR试剂盒,购自深圳匹基公司;Taq DNA聚合酶、dNTP、RNA酶抑制剂,购自宝生物工程(大连)有限公司。引物均由宝生物工程(大连)有限公司合成。TRIZol LS、DTT、RNaseOUT、鼠源反转录酶(MLV),购自Invitrogen公司。
1.3 引物设计 参照文献[7]设计合成F基因引物,上游引物为F1:5'-ATGGGCTCCAAACCTTC⁃TAC-3';下游引物F2:5'-TTGTAGTGGCTCTCATC-3',F基因预期片段大小为1700 bp。参照文献[8]设计合成HN基因引物,上游引物HN1:5'-GAT⁃CAGATGAGAGCCACTA CAA-3';下游引物 HN2:5'-GATAGATGTGACTCTGGTAGGAT-3'。HN基因预期片段大小为2267 bp。新城疫通用反转录引物:5'-ACGGGTAG AA-3'。
1.4 病毒的检测鉴定 病料来源于本中心对辽宁省禽屠宰场的例行监测任务。研磨样品并提取核酸,应用NDV核酸荧光RT-PCR试剂盒检测。将4份阳性病料研磨液接种SPF鸡胚,收集尿囊液[8],按照ND诊断技术标准进行HA和HI试验[9],尿囊液均能被ND标准血清抑制,但不能被H5、H7、H9亚型AIV标准血清和EDS-76血清抑制,分别命名为 CK/CY1/2013、CK/DD2/2013、CK/YK1/2013、CK/TL4/2013,分别简称为CY1株、DD2株、YK1株和TL4株。
1.5 F基因和HN基因的扩增及序列测定 取各分离株的尿囊液200μL,按照Invitrogen公司TRIZol说明书操作,提取RNA。反转录反应体系:RNA悬液2μL,20 pmolRT反应引物2μL,混匀,70℃水浴5 min后立即冰浴3 min,依次加入dNTP(2.5 mmol/μL)4μL,0.1mol/L DTT 4μL,M-MLV 1μL(10 U),RNA酶抑制剂1μL(20 U),5×反转录酶缓冲液8μL混合均匀后。37℃水浴3 h。所获得cDNA用于F、HN基因片段扩增,PCR扩增体系:2.5μL 10×Taq DNA聚合酶缓冲液,0.5μL dNTPs,0.5μL Taq DNA聚合酶(5U),上、下游引物各0.5 μL,1.5μL cDNA,加灭菌超纯水至25μL。PCR反应条件:95℃5 min;94℃2 min、53℃1 min、72℃2 min,进行35个循环;72℃延伸10 min。取5μL PCR产物进行1%琼脂糖凝胶电泳。阳性PCR产物送宝生物工程(大连)有限公司测序。
1.6 F基因、HN基因系统进化树及序列相似性分析 用Lasergene7.0的MegAlign对从GenBank下载32株NDV的F基因序列和19株NDV的HN基因序列进行序列分析。
1.7 致死鸡胚平均时间(MDT)测定 按照《中华人民共和国兽用生物制品质量标准(二〇〇一年版)[10]规定执行。
1.8 F基因的生物信息学分析 利用Lasergene 7.0软件和ExPASy服务器上的Prot Param软件(http://web.expasy.org/protparam/),分析NDV F基因的分子理化性质;利用TMHMM Server v.2.0软件(http://www.cbs.dtu.dk/services/TMHMM-2.0/)分析F蛋白的序列跨膜区;利用SignalP 4.1软件(http://www.cbs.dtu.dk/services/SignalP/)预测分析F蛋白的信号肽[11]。
2 结果
2.1 病毒检测及鉴定结果 所接SPF鸡胚均在72 h以内死亡,死亡鸡胚全身水肿、充血。收获尿囊液进行HA和HI试验。HA和HI试验结果表明,4个分离株的尿囊液均可凝集鸡红细胞,HA凝集效价均在27以上,不能被禽流感阳性血清所抑制,初步判定为NDV。ND病毒核酸荧光RT-PCR检测均为阳性,采用F、HN基因PCR引物均能扩增出相应条带。
2.2 F基因系统进化树及序列相似性分析结果 应用Lasergene 7.0软件的MegAlign软件Clust⁃al W方法构建NDV F基因的系统发生树,将NDV划分为9个基因型 (图1)。DD2株、YK1株和TL4株的F0蛋白裂解位点序列均为112GRQGRL117,具有弱毒株裂解位点特征[12]。CY1株的F0蛋白裂解位点序列为112RRQKRF117,具有强毒株裂解位点特征[12]。
核苷酸同源性分析结果表明,4个分离株的F基因与基因Ⅰ型代表株Queensland V4、Ⅱ型代表株LaSota、Ⅲ型代表株Mukteswar及Ⅸ型标准强毒F48E9的F基因的同源性分别为96.6%~97.3%、95.9%~99.9%、95.8%~96.2%、96.6%~97.1%。CY1株与基因Ⅶ型代表毒株js02株F基因的核苷酸同源性最高,为98.9%;与疫苗株LaSota核苷酸同源性为95.9%;与疫苗株Queensland V4间的核苷酸同源性96.6%。DD2株、YK1株和TL4株与LaSota株核苷酸同源性均为99.9%,与Queensland V4株核苷酸同源性分别为96.6%、97.3%、97.3%。
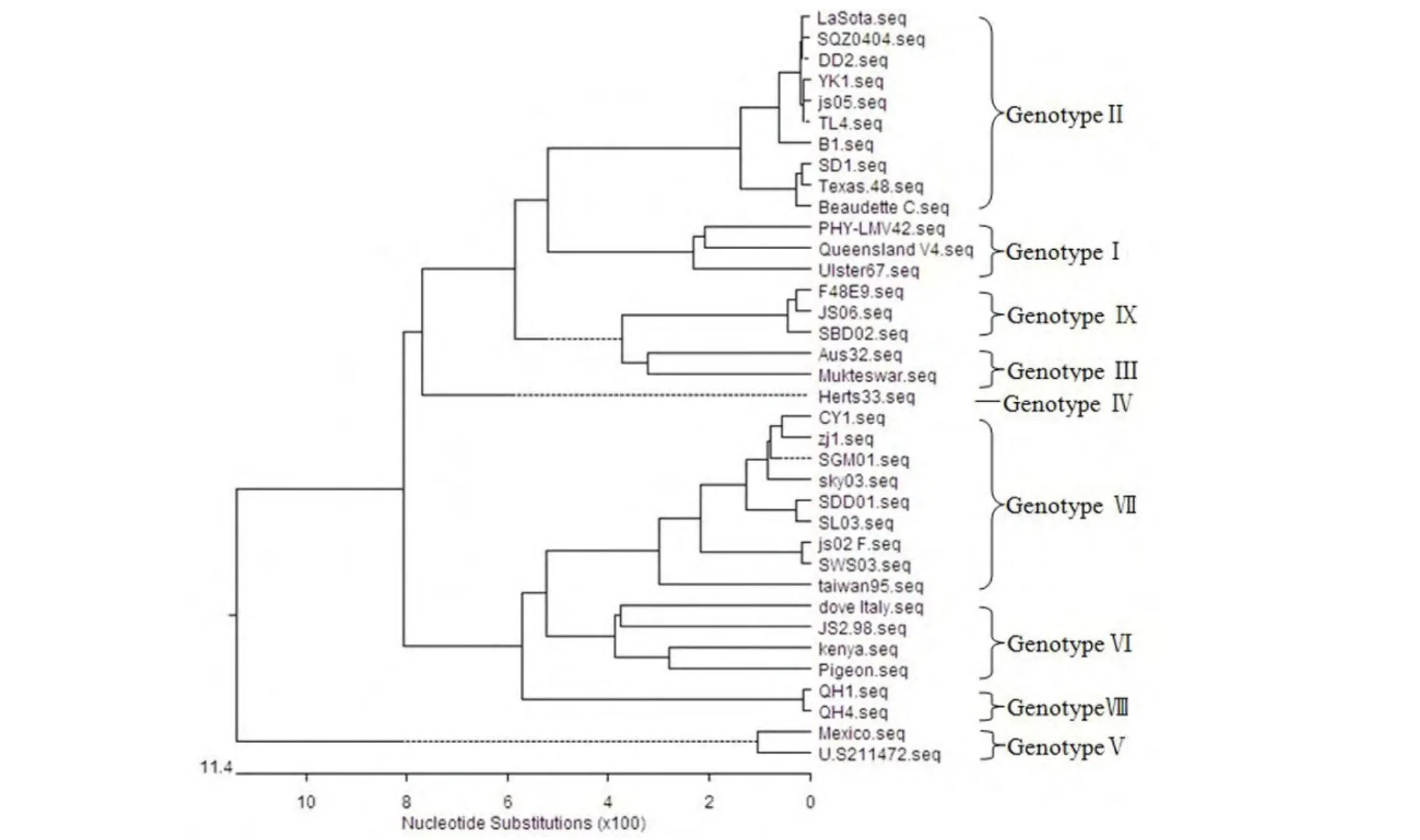
图1 不同ND V毒株F基因进化树
2.3 HN基因系统进化树及序列相似性分析结果 以HN基因序列绘制的进化树显示,将NDV划分为9个基因型(图2)。4个分离株的HN基因之间,DD2株和TL4株间同源性最高,为97.5%;DD2株与CY1株间同源性最低,仅为89%。分离株CY1株与Ⅶ型js02株亲缘关系最近,HN基因同源性为 99.5%;CY1 株与 F48E9、Mukteswar、LaSota、Queensland V4间HN基因的核苷酸同源性分别为94.4%、90.6%、89.0%、83.7%。分离株DD2株、YK1株和TL4株与LaSota株HN基因之间同源性分别为99.9%、96.8%、97.5%;与Queensland V4株HN基因同源性均为92.2%、89.1%、95.1%;与Mukteswar株HN基因之间同源性分别为92.8%、89.7%、91.7%;与F48E9株HN基因之间同源性分别为91.9%、88.7%、94.4%。
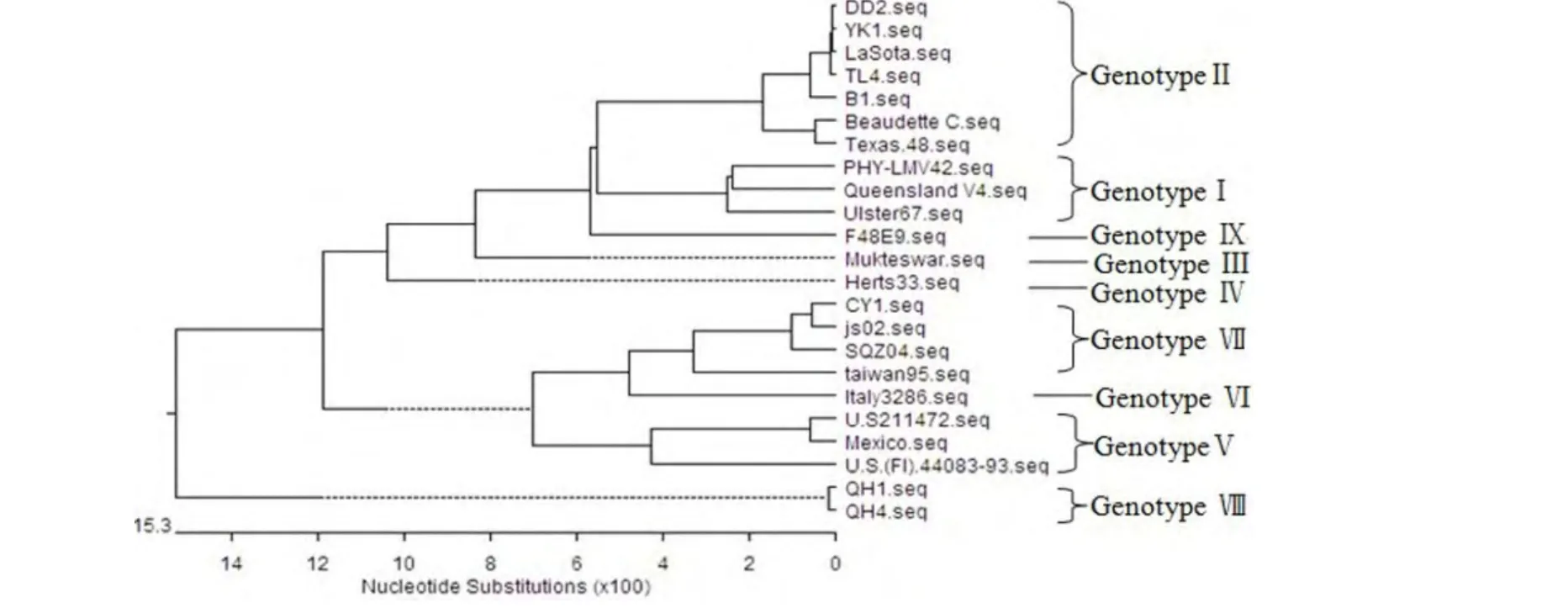
图2 不同ND V毒株HN基因进化树

表1 4个ND V分离株的F蛋白生物信息学分析
2.4 致死鸡胚平均时间(MDT)测定结果 CY1株、DD2株、YK1株和TL4株的MDT分别为50.6 h、76.3 h、62.9 h和81.3 h。
2.5 4个NDV分离株的F基因生物信息学分析结果 采用Prot Param软件分析各分离株F基因的理化性质,结果如表1。应用TMHMM Server V2.0软件预测分析表明,各分离株F蛋白质序列均存在2个跨膜区。Signal P软件预测分析表明,各分离株F蛋白第1~31为氨基酸残基为信号肽,第32为氨基酸残基的综合剪切位点分值最高,据此推断各分离株F蛋白均含有信号肽,均为分泌性蛋白,最大可能剪切位点在第31~32位氨基酸之间。
3 讨论
F蛋白裂解位点是NDV毒力的主要决定因素,但因长期大剂量、高频次ND疫苗免疫所形成的高免疫压力及RNA病毒自身易变异特性,NDV易发生点突变,因此,NDV的分子流行病学研究首选对F基因进行分析,亦有大量研究对HN基因进行分析。在本研究中,采用HN基因序列进行基因型划分的结果(图2),与采用F基因序列进行基因型划分的结果相同(图1),这也与相关研究结果相似[7]。通过裂解位点分析及与疫苗株LaSota、Queensland V4间亲缘关系分析,表明DD2株、YK1株和TL4株属于基因Ⅱ型,与LaSota株的亲缘关系最近,与Queensland V4株亲缘关系均较远,这可能与LaSota株疫苗普遍使用有关,但是否为疫苗突变株值得进一步研究。而CY1株属于Ⅶ型,与目前在养禽业中流行的Ⅶ型js02株的亲缘关系非常近,与LaSota、Queensland V4亲缘关系均较远。
根据毒力不同,NDV可分为强毒株、中强毒株及弱毒株3类;依据兽用生物制品规程规定,MDT小于60 h为强毒,在60~90 h为中强毒,大于90 h为弱毒;依据致病性分子基础的F蛋白裂解位点的不同进行分类,中强毒株、强毒株裂解位点氨基酸序列多为112R/K-R-Q/K-K/R-R-F117,弱毒株裂解位点氨基酸序列则多为112G/E-K/R-Q-G/E-RL117[12]。CY1株的F蛋白裂解位点及其MDT,表明其具有强毒株的特征,同时CY1株相关特性与从本省分离到的3个Ⅶ型毒株强毒株阜新株、沙河株、盘锦株的特性相似[13],DD2株、YK1株和TL4株的MDT均在60~90 h,可归类于中等毒力范围内,裂解位点具有非强毒株的特征,因此还需要进行4个分离株的ICPI和IVPI试验以进一步研究。鉴于F蛋白可介导病毒囊膜与靶细胞膜的融合,促使病毒基因组进入胞浆,为感染性病毒粒子侵入宿主细胞的必需结构,因此本研究采用生物信息学技术对理化性质、跨膜区及信号肽进行初步分析,为进一步研究奠定基础。
[1]Czegledi A,Ujvari D,Somogyi E,et a1.Third genome size cate⁃gory of avian paramyxovirus serotype I(Newcastle disease virus)and evolutionary implications[J].Virus Res,2006,120(1-2):36-48.
[2]Miller P J,Decanini E L,Afonso C L.Newcastle disease:evolu⁃tion of genotypes and the related diagnostic challenges[J].Infect Genet Evol,2010,10:26-35.
[3]Liang R,Cao D J,Li J Q,et a1.Newcastle disease outbreaks in western China were caused by the genotypes VIIa and VIII[J].Vet Microbiol,2002,87:193-203.
[4]Bagai S,Lamb R A.Quantitative measurement of paramyxovirus fusion:differences in requirements of glycoproteins between sim⁃ian virus 5 and human parainfluenza virus 3 or Newcastle dis⁃ease virus[J].JVirol,1995,69(11):6712-6719.
[5]Peeters B P,de Leeuw O S,Verstegen I,et a1.Generation ofa re⁃combinant chimeric Newcastle disease virus vaccine that allows serological differentiation between vaccinated and infected ani⁃mals[J].Vaccine,2001,19(13-14):1616-1627.
[6]秦卓明,马保臣,何叶峰,等.新城疫病毒HN和F基因遗传变异相关性的研究[J].微生物学报,2006,46(2):227-232.
[7]吴双,黄伟平,王伟伟,等.2008年江苏地区基因VIId亚型新城疫病毒遗传变异分析[J].畜牧兽医学报,2009,40(12):1782-1787.
[8]殷震,刘景华.动物病毒学[M].2版,北京:科学出版社,1997:746-747.
[9]刘华雷,吴艳涛,王志亮,等.中华人民共和国国家标准,新城疫诊断技术[S].GB/T 16550-2008.
[10]农业部.中华人民共和国兽用生物制品质量标准(二〇〇一年版)[S].北京:中国农业科技出版社,2001:316-318.
[11]刘建,汤德元,罗险峰,等.猪细小病毒非结构蛋白NS1基因的克隆、序列分析及蛋白质结构预测[J].中国畜牧兽医,2013,40(5):8-13.
[12]Glickman R L,Syddall R J,Iorio R M,et al.Quantitative basic residue requirement in the cleavage activation site of the fusion glycoprotein as a determinant of virulence for NDV[J].Journal of Virology,1998,62(1):354-356.
[13]董国英,徐连壁,顾贵波,等.新城疫F-Ⅶ病毒辽宁强毒株的分离鉴定及防控对策[J].现代畜禽养殖业,2003(6):56-59.
猜你喜欢
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
家禽科学(2019年2期)2019-06-11
食品与生活(2018年7期)2018-09-19
中国畜禽种业(2018年10期)2018-01-19
中国猪业(2017年11期)2017-12-11
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
