
青海撒拉族人群21个短串联重复序列基因座的遗传多态性
2013-11-16 07:02:52王延滨王建文
中国医学科学院学报 2013年5期
马 骏,王延滨,李 开,王建文
1江苏省常州市公安局物证鉴定所,江苏常州213003
2北京市公安局西城分局刑事侦查支队,北京100052
3南京医科大学法医学系,南京210029
短串联重复序列 (short tandem repeat,STR)是广泛存在于人类基因组中的一类遗传标记,其核心重复序列为2~6 bp[1],具有高度多态性,是群体遗传学、疾病相关基因筛选、法医学个体识别等研究的重要工具[2]。本研究对青海省化隆县甘都镇撒拉族人群在D1GATA113、D1S1627、 D1S1677、D2S441、 D2S1776、D3S4529、 D4S2408、 D5S2500、 D6S474、 D6S1017、D9S1122、D10S1248、D10S1435、D11S4463、D12ATA63、D14S1434、 D17S1301、 D18S853、 D19S433、 D20S482、D22S1045共21个STR基因座的遗传多态性进行了调查,获得其等位基因频率分布,并与浙江汉族[3]、宁夏汉族[4]、拉萨藏族[5]、湖北土家族[6]人群的遗传学差异进行统计学分析,旨在丰富我国少数民族人群STR基因座的遗传资料数据库,同时为群体遗传学、疾病相关基因筛选、法医学个体识别等研究提供基础数据。
材料和方法
样本采集 按照“知情同意和知情选择”的原则,采集青海省化隆县甘都镇120名撒拉族无关健康个体 (3代之内均居住于该地区)的静脉血3 ml,EDTA抗凝,-70℃保存备用。
主要仪器和试剂 鼎永4800型扩增仪 (北京鼎永泰克科技有限公司)、GeneAmp PCR System 9700扩增仪 (美国PE公司)、ABI PRISM 3130XL遗传分析仪 (美国 AB公司)、5%Chelex-100(德国默克公司)、AGCU21+1试剂盒 (江苏无锡中德美联生物技术有限公司)。
基因组DNA提取 参照 Walsh等[7]的方法,用5%Chelex-100提取120名撒拉族个体的静脉血基因组DNA。
21个STR基因座的复合扩增 参照AGCU21+1试剂盒说明,扩增的总反应体系为25 μl,其中含预混合液 10 μl,引物 5 μl,热启动 G-Taq 酶 0.5 μl,模板DNA 1 μl,超纯水补足体系到 25 μl。扩增反应在GeneAmp PCR System 9700扩增仪上进行,95℃预变性11 min;94℃ 1 min,62℃ 1 min,72℃ 1 min,共10个循环;90℃ 1 min,60℃ 1 min,72℃ 1 min,共20个循环;60℃延伸60 min,扩增产物4℃保存备用。
扩增产物的毛细管电泳分离及检测 取12 μl去离子甲酰胺、0.5 μl AGCU21+1 Marker SIZ-500与1 μl扩增产物混匀,95℃变性3 min后迅速置于冰上。然后用ABI PRISM 3130XL遗传分析仪对变性后扩增产物进行自动毛细管电泳分离和检测,DNA Collection 2.1软件收集电泳数据,Gene-Mapper 3.2软件分析基因产物片段长度并对等位基因进行分型。
统计学处理 应用Powerstats软件 (http://www.promega.com/geneticidtools/powerstats)计算21个STR基因座的等位基因频率与基因型频率,并应用χ2检验对基因型分布进行 Hardy-Weinberg平衡分析,同时计算个体识别能力 (power of discrimination,DP)、多态信息含量 (polymorphism information contents,PIC)、杂和度 (heterozygosity,H)、非父排除率 (power of exclusion,PE)等群体遗传学指标[8-11]。在获得上述数据的基础上应用SPSS 13.0软件对本研究的撒拉族人群分别与浙江汉族、宁夏汉族、拉萨藏族、湖北土家族人群在21个STR基因座上的等位基因频率分布差异进行统计分析,P<0.05为差异具有统计学意义。
结 果
等位基因频率与基因型频率分布 120名撒拉族个体在21个STR基因座的等位基因频率分布见表1。21个STR基因座共检测到151个等位基因,频率分布在0.0042~0.4917,基因型共有371种,频率分布在0.0083~0.3750(表2)。经 χ2检验,基因型频率分布差异无统计学意义 (P>0.05),符合Hardy-Weinberg平衡定律。

表1 青海省化隆县甘都镇撒拉族人群在21个STR基因座的等位基因频率 (n=120)Table 1 Allele frequencies of 21 STR loci of Salar minority ethnic group from Gandu town,Hualong county of Qinghai province(n=120)

(续表1)
21个STR基因座的群体遗传学指标 120名撒拉族个体在D1GATA113、D1S1627、D1S1677、D2S441、D2S1776、 D3S4529、 D4S2408、 D5S2500、 D6S474、D6S1017、D9S1122、D10S1248、D10S1435、D11S4463、D12ATA63、D14S1434、D17S1301、D18S853、D19S433、D20S482、D22S1045基因座的DP、H、PIC、PE等多态性指标见表2。DP值介于0.796~0.948,H值介于0.650~0.817,PIC值介于0.590~0.810,PE值介于0.355~0.630。累积耦合概率为1.75×10-20,累积非父排除概率为0.9999999(表2)。
撒拉族人群与浙江汉族、宁夏汉族、拉萨藏族、湖南土家族人群的等位基因频率分布差异 在21个STR基因座中,撒拉族人群与浙江汉族人群在14个基因座差异具有统计学意义 (P<0.05),与宁夏汉族人群在12个基因座差异具有统计学意义 (P<0.05),与拉萨藏族人群在12个基因座差异具有统计学意义 (P<0.05),与湖北土家族人群在13个基因座差异具有统计学意义 (P<0.05)(表3)。
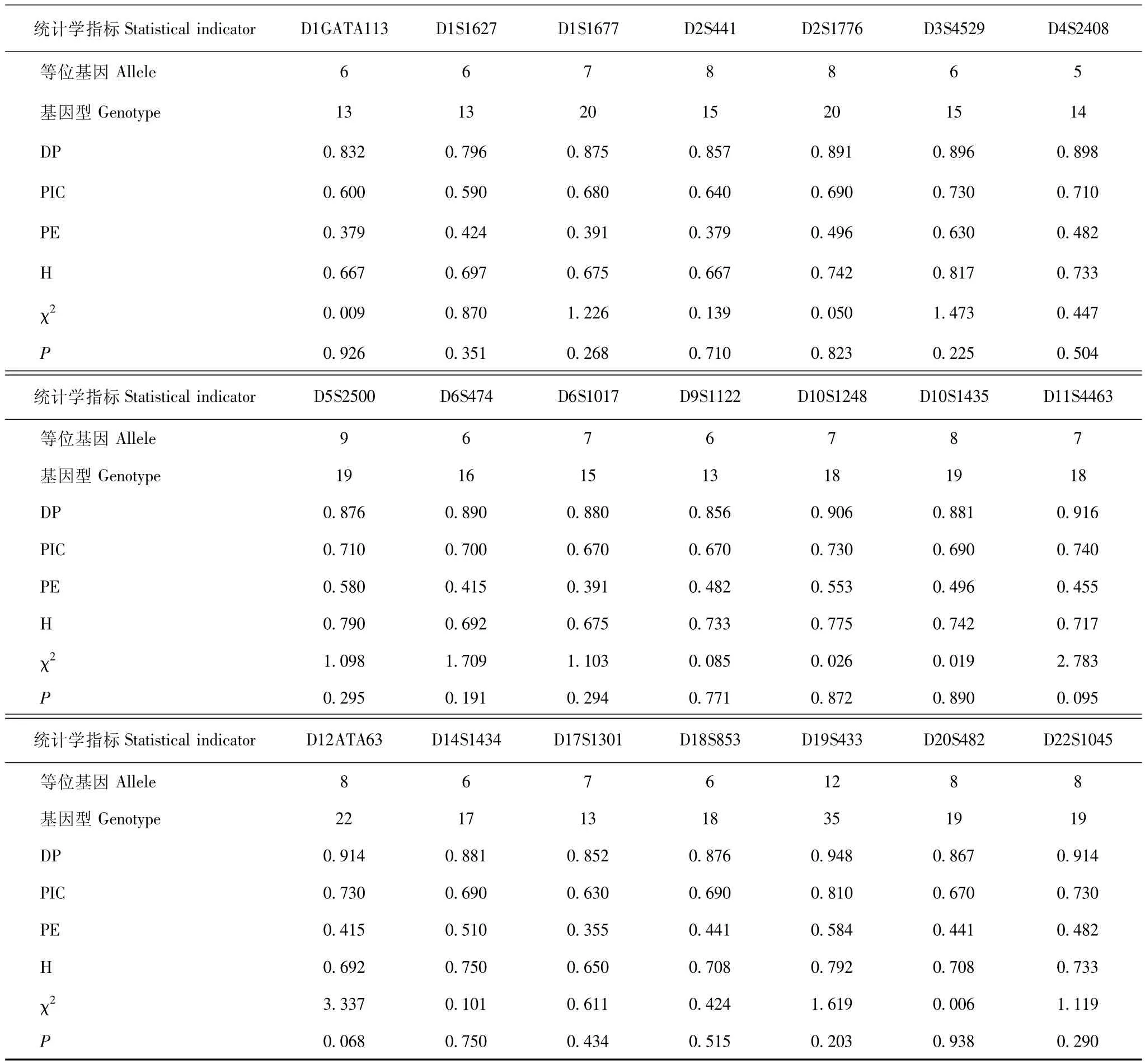
表2 青海省化隆县甘都镇撒拉族人群在21个STR基因座的多态性统计学指标 (n=120)Table 2 Statistical indicators of the polymorphisms of 21 STR loci of Salar ethnic group from Gandu town,Hualong country of Qinghai province(n=120)
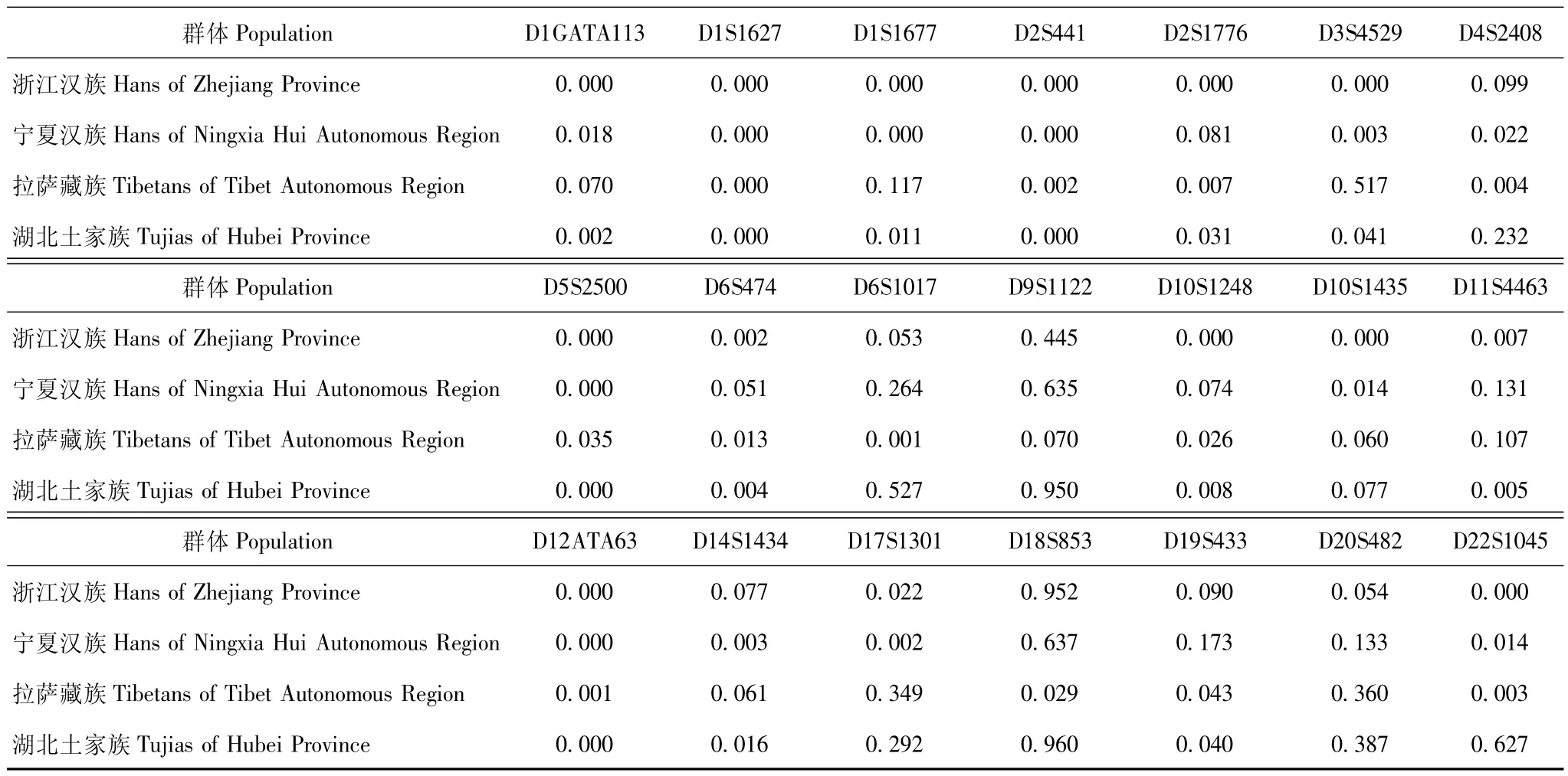
表3 青海省化隆县甘都镇撒拉族人群与其他民族人群在21个STR基因座的等位基因频率分布差异分析 (P)Table 3 Allele frequencies of 21 STR loci between Salar and other ethnic groups in China(P)
讨 论
中国人不同群体之间的遗传差异主要有两类形成原因。一类是民族内部的遗传多态现象,另一类是民族之间的遗传差别[12]。基因组DNA多态性研究突破了古生物学家、历史学家、语言学家以及考古学家研究现代人类群体间关系的局限性。其中,STR基因座的应用最为普遍,其由于核心单位重复数目的变化而在群体中呈现出遗传多态性,反映出群体的分化过程,由此成为人类基因组扫描的首选多态性基因座[13]。通常认为,在一个群体中频率最高的基因座的等位基因是该群体中最原始、最保守的,而其余的等位基因均是在群体分化的过程中由频率最高的等位基因突变形成的[14]。STR基因座在群体之间和群体内部的异质性水平较高,因此不同群体以及亲缘关系相近的群体之间的等位基因和基因型频率分布有明显的差异,并且地理因素在遗传多态性形成中的作用比语言等因素更为显著[15]。因此,在将STR基因座应用于群体遗传学等研究时,必须先对特定的群体进行多态性调查,获得其等位基因频率分布数据。笔者曾对西安地区汉族人群、拉萨地区藏族人群进行过STR基因座的群体学调查研究[16-17]。
撒拉族主要聚居在青海省循化撒拉族自治县和化隆回族自治县,化隆县境内的撒拉族均是由循化地区陆续迁移而来,约有64%的撒拉族聚居于甘都镇。Teng等[18]已经对循化地区的撒拉族人群进行过21个STR基因座多态性研究,本研究调查21个STR基因座在青海省化隆县甘都镇120名撒拉族个体中的多态性分布,得到了反映该地区遗传特征的等位基因频率和基因型频率数据。本研究共检测出151个等位基因与371种基因型,DP值介于0.796~0.948,PE值介于0.355~0.630,累积耦合概率为1.75×10-20,累积非父排除概率为0.9999999。在Teng等[18]对青海循化地区的撒拉族人群调查中,累积耦合概率为8.26×10-20,累积非父排除概率为 0.999999,研究结果一致。
STR基因座的多态性及其应用价值一般用DP、H、PIC、PE衡量。Shriver等[19]认为,PIC值>0.5时表明该遗传标记可提供高信息量,并且当该遗传标记的DP≥0.8或者PE≥0.5时,该遗传标记具有高度多态性。据此标准,本研究中的D1GATA113、D1S1677、D2S441、 D2S1776、 D3S4529、 D4S2408、 D5S2500、D6S474、D6S1017、D9S1122、D10S1248、D10S1435、D11S4463、 D12ATA63、D14S1434、D17S1301、D18S853、D19S433、D20S482、D22S1045共20个STR基因座在撒拉族人群中均属于高度多态性基因座。研究结果同时显示,D1S1627基因座的 DP值、H值均为最低(分别为0.796、0.590),与邵伟波等[20]在华东汉族人群中的研究一致,属于中度多态性基因座。上述21个STR基因座联合应用完全能满足撒拉族人群的群体遗传学研究和法医学个体识别的需要。
将本研究中的撒拉族人群分别与浙江汉族、宁夏汉族、拉萨藏族、湖北土家族进行统计学分析,结果显示分别在14、12、12、13个STR基因座差异具有统计学意义。分析其原因,浙江汉族与宁夏汉族可能是由于地理因素的原因而造成了与撒拉族差异程度不同。而拉萨藏族、湖北土家族与撒拉族的差异则可能是由于民族因素、地理因素共同作用造成的。另外,部分基因座差异并无统计学意义,分析可能有以下两个原因:(1)人群之间在此类基因座确实不存在差异;(2)研究的样本量偏小,使得此类基因座的多态性未充分显示。
综上,本研究调查的21个STR基因座在青海省化隆县甘都镇撒拉族人群中联合应用具有良好的遗传多态性。同时,由于STR基因座在不同民族、地域之间的人群中的差异,在进行群体遗传学、疾病相关基因筛选、法医学个体识别等研究时,应采用本地基础数据,并且尽可能选择更适合本地人群的STR基因座,同时也应在现有的基础上增加样本量,由此得出的分析结果其可信度将会更高。
[1]Edwards A,Civitello A,Hammond HA,et al.DNA typing and genetic mapping with trimeric and tetrameric tandem repeats[J].Am J Hum Genet,1991,49(4):746-756.
[2]Sprecher CJ,Puers C,Lins AM,et al.General approach to analysis of polymorphic short tandem repeat loci[J].Biotechniques,1996,20(2):266-271.
[3]吴微微,徐志成,郝宏蕾,等.浙江汉族人群21个非CODIS系统STR基因座的遗传多态性调查 [J].刑事技术,2010,3:19-22.
[4]Shen CM,Wang HD,Liu WJ,et al.Allelic diversity distributions of 21 autosomal short tandem repeat loci in Chinese Ningxia Han population [J].Forensic Sci Int Gene,2013,7(3):e78-e79.
[5]Zhu BF,Shen CM,Wang HD,et al.Genetic diversities of 21 non-CODIS autosomal STRs of a Chinese Tibetan ethnic minority group in Lhasa [J].Int J Legal Med,2011,125(4):581-585.
[6]Yuan GL,Shen CM,Wang HD,et al.Genetic data provided by 21 autosomal STR loci from Chinese Tujia ethnic group[J].Mol Biol Rep,2012,39(12):10265-10271.
[7]Walsh PS,Metzger DA,Higuchi R.Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic materia1 [J].Biotechniques,1991,10(4):506-513.
[8]Nei M.Estimation of average heterozygosity and genetic distance from a small number of individual[J].Genetics,1978,89(3):583-590.
[9]Jones DA.Blood samples:probabilities and discrimination[J].J Forensic Sci Soc,1972,12(2):355-359.
[10]郑秀芬.法医DNA分析[M].北京:中国人民公安大学出版社,2002:405-421.
[11]Ohno Y,Sebetan IM,Akaishi S.A simple method for calculating the probability of excluding paternity with any number of codominant alleles [J].Forensic Sci Int,1982,19(1):93-98.
[12]翁自立,袁义达,杜若甫.中国14个民族红细胞血型座位的遗传分化 [J].遗传学报,1990,17(4):260-268.
[13]Hammond HA,Jin L,Zhong Y,et al.Evaluation of 13 short tandem repeat loci for use in personal identification application[J].Am J Hum Genet,1994,55(1):175-189.
[14]Li SL,Yamamoto T,Yoshimoto T,et al.Phylogenetic relationship of the population within and around Japan using 105 short tandem repeat polymorphic loci[J].Hum Genet,2006,118(6):695-707.
[15]Rosser ZH,Zerjal T,Hurles ME,et al.Y-chromosomal diversity in Europe is clinal and influenced primarily by geography,rather than by language [J].Am J Hum Genet,2000,67(6):1526-1543.
[16]王振原,马骏,徐永城,等.西安地区汉族人群13个STR基因座多态性分析 [J].中南大学学报,2005,30(2):236-238.
[17]马骏,巴华杰,张文杰,等.拉萨地区藏族人群17个短串联重复序列基因座的遗传多态性[J].中国医学科学院学报,2011,33(4):397-401.
[18]Teng Y,Zhang FX,Shen CM,et al.Genetic variation of new 21 autosomal short tandem repeat loci in a Chinese Salar ethnic group [J].Mol Biol Rep,2012,39(2):1465-1470.
[19]Shriver MD,Jin L,Boerwinkle E,et al.A novel measure of genetic distance for highly polymorphic tandem repeat loci[J].Mol Biol Evol,1995,12(5):914-920.
[20]邵伟波,张素华,李莉.21个非CODIS STR基因座的遗传多态性 [J].法医学杂志,2011,27(1):36-38.
猜你喜欢
智慧健康(2021年17期)2021-07-30 14:38:32
心声歌刊(2018年4期)2018-09-26 06:54:14
现代检验医学杂志(2016年5期)2016-08-20 03:16:54
法医学杂志(2015年4期)2016-01-06 12:36:40
法医学杂志(2015年4期)2016-01-06 12:36:38
食管疾病(2015年3期)2015-12-05 01:45:11
群文天地(2015年2期)2015-04-16 01:02:35
分子诊断与治疗杂志(2013年3期)2013-07-08 02:17:11
中国刑警学院学报(2012年1期)2012-04-24 10:38:00
黑龙江史志(2011年20期)2011-08-15 00:46:01
