
康复训练对脑梗死大鼠认知功能、海马内突触素和神经颗粒素表达的影响
2012-08-09 03:50李雨峰吴莹程明徐丽黄林李怡余茜
中国康复理论与实践 2012年1期
李雨峰,吴莹,程明,徐丽,黄林,李怡,余茜
脑梗死是一种严重危害人类健康的常见病、多发病,幸存者中约70%~80%有不同程度的后遗症,40%不能生活自理。这些存活的患者常遗留瘫痪、失语等严重残疾并有相应的认知障碍,其中以学习记忆障碍最为常见、持久。如何促进脑梗死后学习记忆功能的恢复成为现代康复治疗研究的重点。
大量研究提示,运动训练能通过促进中枢神经系统的神经再生、修复和突触的可塑性而改善机体的功能。突触素(synaptophysin,Syn)为突触前膜标志蛋白,是一种与突触结构和功能极为相关的糖蛋白,几乎所有中枢和周围神经系统的突触前终末内均发现有突触素[1]。神经颗粒素(neurogranin,Ng)是一种新发现的脑特异性突触后蛋白,主要在大脑皮质、海马、纹状体和杏仁核中高表达,可能在学习、记忆、神经系统发育(突触可塑性)等生理过程中发挥重要作用[2-5]。本研究采用线栓法制成大鼠右侧大脑中动脉缺血梗死模型,并对康复组大鼠进行训练,采用免疫组织化学技术观察突触前膜标志蛋白突触素及突触后蛋白神经颗粒素的表达变化,并进行神经功能及运动功能评定,探讨运动训练对脑梗死大鼠认知功能影响的可能机制。
1 材料与方法
1.1 实验动物 健康雄性Wistar大鼠75只,体重(250±50)g,8周龄,由泸州医学院实验动物科提供。
1.2 试剂和仪器 试剂:多聚甲醛;戊巴比妥钠;Syn免疫组织化学试剂盒;Ng免疫组织化学试剂盒;多聚赖氨酸;DAB显色试剂盒(棕色)等。仪器:Ax70荧光显微镜;DP-70图像分析系统;Y-型迷宫;85-2型磁力搅拌器等以及自制滚筒网状训练仪、平衡训练棒、转棒训练仪、网屏训练仪。
1.3 方法
1.3.1 模型建立及分组 参照小泉线栓法[6]制成右侧大脑中动脉缺血梗死模型。手术后2 h符合以下4项体征即模型成功:①动物苏醒后表现为提尾时左侧前肢内收屈曲;②同侧Hornor氏征;③爬行时向左划圈;④站立时左侧倾倒。
将造模成功的48只动物随机分为脑梗死自由活动组(模型组,n=24)和脑梗死运动训练组(康复组,n=24),每组又分为术后15 d、25 d、35 d 3个亚组,每个亚组各8只。各组大鼠都处于相同的环境喂养中。
1.3.2 运动训练 康复组大鼠在手术后第5天开始进行运动训练,采用以下装置:滚筒网状式训练仪:训练大鼠抓握、旋转及行走能力;平衡训练:训练其平衡能力;网屏训练:训练大鼠的抓握能力及肌力;转棒训练:训练其动态平衡。40 min/d,6d/周。
1.3.3 神经功能和运动功能评定 两组分别于术后15d、25d、35d进行神经功能评定和运动能力评分。
神经功能评分采用Bederson等[7]的评定方法:0级:未有神经缺失症状;1级:将大鼠尾巴提起,瘫痪侧前肢回收屈曲于腹下,正常侧前肢向地面伸展;2级:除1级体征外,向瘫痪侧推大鼠时阻力较对侧明显降低;3级:除以上体征外,大鼠有向瘫痪侧旋转的行为。
平衡木行走测评:0分:能跳上平衡木,在上面行走不会跌到;1分:能跳上平衡木,在上面跌到的机会少于50%;2分:能跳上平衡木,跌倒的机会大于50%;3分:在健侧后肢帮助下能跳上平衡木,但瘫痪侧后肢不能帮助向前移动;4分:在平衡木上不能行走,但可坐在上面;5分:将鼠放在平衡木上会掉下来。
网屏实验:0分:前爪握住网屏大约5s不会掉下来;1分:暂时握住网屏,需滑落一段距离,但没有掉下来;2分:在5s内掉下来;3分:网屏转动时,鼠即刻掉下来。
转棒行走测评:0分:转动过程中,鼠可在棒上面行走;1分:在转动过程中,鼠不会掉下来,时间60s以上;2分:转动开始后鼠从棒上掉下来;3分:转动开始前,鼠就从棒上掉下来。
1.3.4 学习记忆能力 术后35 d对相应亚组大鼠进行Y-型迷宫[8]测试:实验前,将大鼠放入迷宫,使其适应5 min,然后开始实验。大鼠在通电后从所在亮臂跑到另一亮臂记为错误,跑到暗臂记为正确。每天训练30 min,连续刺激10次后大鼠休息2 min,记录大鼠学会(连续10次训练中有9次正确即学会)所需的训练次数,训练次数越少表明大鼠学习能力越强。
1.3.5 取材及免疫组化 所有大鼠在康复训练后各时间点评分后分别取材,每个亚组取材8只。在视交叉前2 mm至视交叉后2 mm范围内(含海马的脑段)冠状取材,经脱水—透明—浸蜡—包埋。进行Syn及Ng免疫组织化学染色。用DP-70图像采集系统采集图片,Image-pro Plus 5.1软件进行半定量分析。每张切片随机取上下左右中5个视野,分别测定Syn阳性产物信号的面积和Ng阳性产物信号的面积占当时视野面积的百分比,测定其阳性产物平均光密度(OD)值,并减去背景光密度值,得到校正光密度值。
2 结果
在造模程中,模型不成功或死亡27只,分组后,模型组2只、康复组3只大鼠死亡,均进行及时随机补充,保证每组大鼠数量不变。
2.1 神经功能与运动功能 康复组神经功能、平衡木行走、网屏实验和转棒行走术后15 d与同期模型组比较无显著性差异(P>0.05),术后25 d、35 d均低于模型组(P<0.05)。见表1~表4。

表1 两组各时间点神经功能测评比较

表2 两组各时间点平衡木行走测评比较

表3 两组各时间点网屏实验测评比较
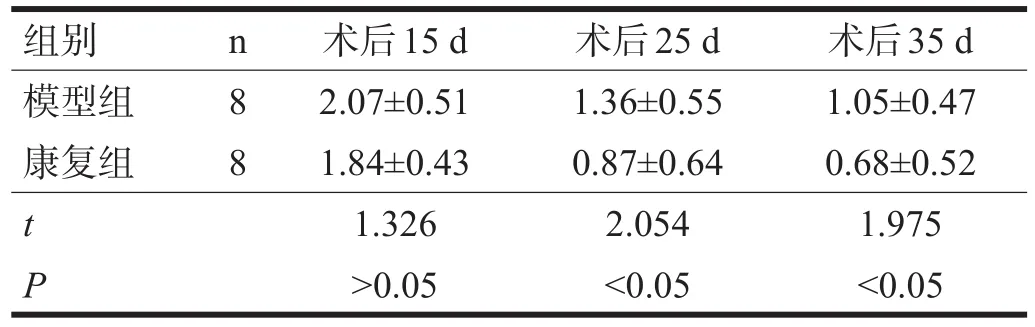
表4 两组各时间点转棒行走测评比较
神经功能评分与转棒评分(r=0.574,P<0.01)、平衡木评测(r=0.685,P<0.01)、网屏评测(r=0.703,P<0.01)呈明显正相关。
2.2 学习记忆能力 康复组大鼠术后35 d Y-迷宫分辨学习能力(67.34±8.05),明显优于模型组(102.58±11.73)(t=1.982,P<0.05)。神经功能与学习记忆能力呈明显正相关(r=0.567,P<0.01)。
2.3 Syn、Ng表达 康复组Syn校正光密度值在术后35 d优于模型组(P<0.05);Ng校正光密度值在术后25 d、35 d均优于模型组(P<0.05)。见表5、表6。
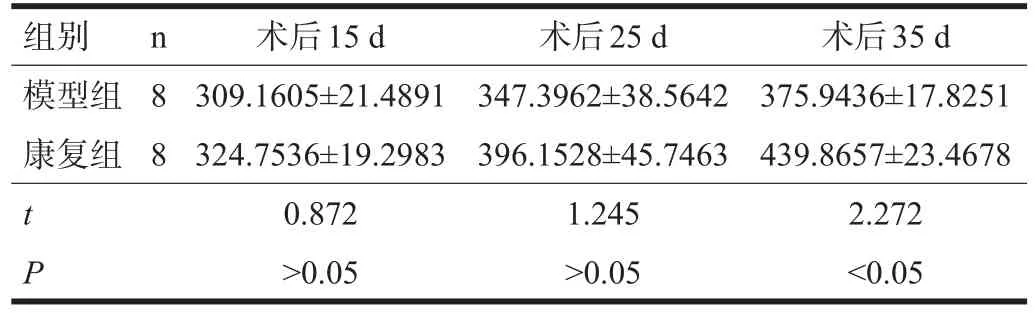
表5 Syn在梗死侧海马CA3区的表达(校正光密度值)

表6 Ng在梗死侧海马齿状回的表达(校正光密度值)
3 讨论
Syn为突触前膜标志蛋白,其密度可间接反映突触的数量、分布和密度[9]。本研究显示,梗死侧Syn阳性反应物主要分布在纹状体、海马及下丘脑。在术后25 d,康复组阳性颗粒数量明显增多,颗粒小而密集,染色均匀,可见明显Syn阳性纤维束;术后35 d康复组Syn阳性纤维排列整齐,较前稍有增多,并可见Syn阳性纤维网,校正光密度与模型组比较有显著性差异(P<0.05)。提示康复训练可能有促进海马突触结构的改建和再建。由于突触素是构成突触囊泡特异性的膜通道,突触素的增加意味着突触囊泡转运能力的增强,传递效能增强,推测康复训练可能通过改善突触的数量或(和)效能来改善认知能力。
Ng广泛分布于大脑皮质、海马、嗅球等重要脑区的神经突触后。目前研究表明,Ng在神经的可塑性及脑部的一些病理生理变化等方面起重要作用,尤其是作为Ca2+敏感性的钙调蛋白(calmodulan,CaM)结合蛋白及蛋白激酶C(protein kinase C,PKC)的突触后底物,参与在学习记忆功能中起核心作用的脑内几种蛋白信号转导途径、长时程增强(1ong-term potentiation,LTP)和长时程抑制(1ong-term depression,LTD)等突触可塑性机制,因而它可能涉及学习记忆的形成和巩固[4,10]。
本研究发现,Ng免疫阳性反应物呈棕色或棕黄色颗粒状沉积,在海马齿状回内呈3条清晰的条带(内、中、外带)。术后15 d康复组和模型组Ng阳性颗粒细小,着色浅淡,沿内带和外带边缘呈条索状不均匀分布;术后25 d康复组阳性颗粒数量增多,着色加深,分布变得密集,从内带和外带逐渐向中带聚集;术后35 d康复组阳性颗粒数量较前明显增多,着色明显加深,颗粒也较前增大。校正光密度在术后25 d康复组较模型组高(P<0.05),术后35 d康复组较模型组明显升高(P<0.01),提示康复训练能够造成与突触可塑性密切相关分子含量的改变,从而引起脑功能的变化。由于Ng是一种不具有酶活性的神经元内蛋白分子,其对突触可塑性的调节与改变细胞内游离Ca2+浓度相关。有文献表明,Ng基因敲除纯合子小鼠表现出严重的空间学习能力障碍。海马脑片中Ca2+-CaM依赖的蛋白激酶Ⅱ(CaMKⅡ)、PKC和cAMP依赖的蛋白激酶的活性在基因敲除纯合子小鼠较正常小鼠显著降低[11]。
认知障碍是脑梗死患者最为常见和持久的并发症,促进脑梗死患者学习记忆能力的改善是脑梗死治疗的重点和难点,目前在中枢可塑性研究中,促进学习记忆能力改善的康复机制备受关注。本实验采用Y-型迷宫评定大鼠逃避条件反射和空间辨别能力,大鼠在Y-型迷宫中学会逃避不适刺激所需的训练次数代表大鼠形成条件反射和整体学习能力,次数越少学习能力越强。康复组和模型组大鼠处死前分别进行Y-迷宫分辨学习结果为术后康复组明显优于模型组(P<0.05),表明康复训练(网状滚筒训练、平衡木训练、转棒训练、网屏训练)对脑梗死大鼠认知能力的恢复有促进作用。脑梗死大鼠神经功能与各项运动功能均呈显著正相关,表明认知能力可随着神经功能和运动能力的恢复而改善。
余茜等发现,运动训练可明显改善运动神经功能和认知能力,使健侧海马突触界面曲率和突触后致密物(PSD)厚度增大,穿孔性突触的百分率增多,突触穿孔后致密物变为节段,出现多个活性区,使得不同受体簇的不同活性区传递功能大大加强,进一步加强了突触传递功能,使其习得性LTP形成速度明显快于模型组[12-15]。并指出运动康复促进脑梗死大鼠认知能力的恢复可能是通过影响脑梗死鼠健侧海马CA3区N-甲基-D-天冬氨酸(N-methyl-D-aspartate,NMDA)受体通道开放导电水平、开放时间和开放概率来实现的。康复训练后健侧海马突触体游离Ca2+浓度变化可影响信息处理过程,最终表现在认知能力行为上。另有研究认为,康复训练促进双侧大脑皮质Fos阳性神经元表达增强[16],认为是细胞长期效应被激活的标志,c-fos基因表达和Fos蛋白的生物合成对促进学习记忆过程至关重要。Tischmeyer等报道,大鼠在完成Y-型迷宫视觉辨别任务后,其海马c-fos mRNA升高,在清醒大鼠海马齿状回诱导LTP可导致c-fos基因表达增高[17],表明康复训练也可通过提高c-fos基因表达来增进认知能力。
总之,运动训练对中枢神经系统的作用可通过增加Syn和Ng阳性表达改善脑梗死大鼠认知能力。
[1]Masliah E,Terry R.The role of synaptic proteins in the pathogenesis of disorders of the central nervous system[J].Brain Patho1,1993,3(1):77-85.
[2]Wu J,Li J,Huang KP,et al.Attenuation of protein kinase C and cAMP-dependent protein kinasse signal transduction in the neuro-granin knockout mouse[J].J Biol Chem,2002,277(22):19498-19505.
[3]Ressler KJ,Paschall G,Zhou XL,et al.Regulation of synaptic plasticity genes during consolidation of fear conditioning[J].J Neurosci,2002,22(18):7892-7902.
[4]van Dam EJ,Ruiter B,Kamal A,et al.N-methyl-D-aspartate-induced long-term depression is associated with a decrease in postsy-naptic protein kinase C substrate phosphorylation in rat hippocampalslices[J].Neurosci Lett,2002,2002(3):129-132.
[5]Krucker T,Siggins GR,McNamara RK,et al.Targeted disruption of RC3 reveals a calmodulin-based mechanism for regulating meta-plasticity in the hippocampus[J].J Neurosci,2002,22(13):5525-5535.
[6]屈秋民,曹振玲,杨剑波.线栓法大鼠大脑中动脉闭塞局灶性脑缺血模型Longa法和小泉法的比较[J].中华神经科杂志,2000,22(5):289.
[7]Bederson JB,Pitts LH,Tiles M,et al.Rat middle cerebral artery occlusion:evaluation of the model and development of a neurologic examination[J].Stroke,1986,17:472-476
[8]赵崇侃,程光,陈启盛.一种智能化的Y迷宫[J].中国应用生理学杂志,1997,13(4):363-365.
[9]Wiedenmann B,Franke WW.Identification and localization of synaptophysin:an integral membrane glycoprotein Mr.38000 characteristic of presynaptic vesicles[J].Cell,1985,41(3):1017-1028.
[10]Ressler KJ,Paschall G,Zhou XL,et al.Regulation of synaptic plasticity genes during consolidation of fear conditioning[J].J Neurosci,2002,22(18):7892-7902.
[11]Huang KP,Huang FL,Jagel T,et al.Neurogranin/RC3 enhances long-term potentiation and learning by promoting calcium-mediated signaling [J].J Neurosci,2004,24(47):l0660-l0669.
[12]余茜,李晓红,吴士明.运动训练后脑缺血大鼠学习记忆与健侧脑内突触结构变化的关系[J].中华物理医学与康复杂志,2002,24(7):399-402.
[13]余茜,李晓红,吴士明,等.运动康复对脑梗死大鼠学习记忆能力和LTP的影响[J].中华物理医学与康复杂志,2002,24(3):140-143.
[14]余茜,李晓红,刘曦,等.康复训练对脑梗死大鼠学习记忆与健侧海马神经元NMDA受体通道的影响[J].中华物理医学与康复杂志,2002,24(11):683-686.
[15]余茜,李晓红,何成松,等.康复训练对脑梗死大鼠学习记忆能力与健侧海马突触体胞浆游离Ca2+浓度的影响[J].中华物理医学与康复杂志,2004,26(4):209-211.
[16]李玲,袁华,徐莉.康复训练对脑梗死大鼠中枢神经系统Fos表达的影响[J].中华物理医学与康复杂志,2001,23(1):7-10.
[17]Tischmeyer W,Grimm R.Activation of immediate early genes and memory formation[J].Cell Mol Life Sci,1999,55(4):564-574.
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05
儿童与健康(幼儿教师参考)(2021年7期)2021-08-03
装备制造技术(2020年11期)2021-01-26
中华养生保健(2020年2期)2020-11-16
快乐作文(3.4年级)(2020年4期)2020-05-25
艺术品鉴(2019年9期)2019-10-16
创新作文(小学版)(2019年10期)2019-09-25
中国生物医学工程学报(2019年6期)2019-07-16
制造业自动化(2017年2期)2017-03-20
饮食科学(2014年1期)2014-02-23
