
烤烟几个重要植物学性状的遗传分析
2012-07-31 07:22张兴伟王志德刘艳华牟建民
中国烟草科学 2012年5期
张兴伟,王志德,任 民,刘艳华,牟建民
(农业部烟草生物学与加工重点实验室,中国农业科学院烟草研究所,青岛 266101)
烟草(Nicotiana tobacumL.)是我国重要的经济作物之一,其中烤烟每年种植面积及产量均占世界三分之一左右。烤烟的植物学性状会直接影响烟叶产值及化学成分,因此受到研究者的关注。国内外多数研究认为,株高、叶数、叶长、叶宽等大多数性状的遗传以加性效应为主,显性效应作用较小[1-6],只有少数性状受显性效应影响较大[4-6],基本无上位性效应[3]。本研究以烤烟品种丸叶和Coker319杂交,获得杂交组合的P1、P2、F1、B1、B2和F2六个世代,以数量性状“主基因+多基因”混合遗传模型进行分析,对重要植物学性状的遗传率和基因效应等参数进行估计,旨在探讨烤烟重要植物学性状的遗传模式,为烤烟株型育种提供理论依据。
1 材料与方法
1.1 试验材料
本试验采用来源于国家烟草中期库的烤烟品种丸叶(P1,引自日本)和Coker319(P2,引自美国)为亲本,构建 F1、F2、B1(F1×P1)和 B2(F1×P2)群体。2009年春季将全部供试材料种植于青岛即墨实验农场,株行距为55 cm×120 cm,亲本和F1每小区种植15株,共45株,B1、B2和F2每小区种植75株,共225株。
1.2 试验方法
1.2.1 性状指标测定 性状指标测定参照文献[7]。
1.2.2 试验设计及数据分析 随机区组设计,3次重复。采用植物数量性状主基因+多基因混合遗传多世代联合分析方法,对该组合6个世代的重要植物学性状进行分析。通过极大似然法和 Iterated expectation and conditional maximization(IECM)估计各世代、各成分的分布参数,然后通过 Akaike′s information criterion(AIC)值选择最佳模型,同时进行一组适合性检验,包括均匀性U12、U22和U32检验,Smirnov 检验(nW2)和 Kolmogorov 检验(Dn),根据检验结果选择最优遗传模型。采用最小二乘法从最优遗传模型的各成分分布参数估计各基因效应值,分析各遗传效应。
在主基因+多基因的混合遗传模型中,表现型值(p)表示为群体平均数(m)、主基因效应(g)、多基因效应(c)和环境效应(e)之和,即 p=m+g+c+e,因此,表现型方差(σ2p)可表示为主基因方差(σ2mg)、多基因方差(σ2pg)和环境方差(σ2e)之和,即 σ2p=σ2mg+σ2pg+σ2e。
通过亲本和 F1同质群体提供环境误差方差的无偏估计,可估计环境方差σ2e,成分分布方差σ2f,群体表型方差 σ2p,进而可得到主基因和多基因的遗传方差(σ2mg、σ2pg)和遗传率(h2mg、h2pg)。

2 结 果
2.1 性状遗传方差分析及世代平均数分析
2个品种间几个植物学性状方差分析表明,重复间差异不显著,处理间差异显著(表1),说明2个亲本重要植物学性状在遗传方面存在着真实稳定的差异,可以作为研究烤烟重要植物学性状遗传规律的材料进行分析。
从表1还可以看出,B1、B2和 F2的株高位于双亲之间,F1超过高值亲本P1,说明株高的遗传表现一定的超显性效应,另外叶宽和节距的遗传也表现一定的超显性效应;叶数、茎围遗传表现一定的负超显性效应,叶面积、比叶重遗传表现一定的显性效应,移栽至中心花开放天数、鲜叶重、叶形指数、叶长及蒴果重遗传表现出一定的隐性效应。
2.2 性状的遗传模型
2.2.1 株高遗传模型 用主基因+多基因混合遗传模型对丸叶×Coker319组合6世代株高进行联合分析,通过IECM算法获得1对主基因(A)、2对主基因(B)、多基因(C)、1对主基因+多基因(D)和2对主基因+多基因(E)共5类24种遗传模型的极大似然函数值和AIC值。根据AIC准则,E0、E1和E3的AIC值较小,作为备选遗传模型。进一步对E0、E1和E3模型进行一组适合性[U12、U22、U32检验,Simirnov(nw2)检验和Kolmogorov检验(Dn)]检验,结果表明,在30个检验统计量中,E0模型没有统计量与该模型差异达到显著水平,E1模型有7个统计量达到显著水平,E3模型有8个统计量达到显著水平,综合确定 E0为株高最优遗传模型,即 2对加性-显性-上位性主基因+加性-显性-上位性多基因模型。

表1 各世代重要植物学性状参数平均值Table 1 Means of the botanic trait parameters among six generations
2.2.2 叶数遗传模型 通过IECM算法获得5类24种遗传模型的极大似然函数值和AIC值。根据AIC准则,B1、E0、E1和E3的AIC值较小,作为备选遗传模型。适合性检验表明,在30个统计量中,E0模型没有统计量达到显著水平,其他几个模型均有统计量达到显著水平,综合确定 E0为叶数最优遗传模型,即 2对加性-显性-上位性主基因+加性-显性-上位性多基因模型。
2.2.3 叶面积遗传模型 根据AIC准则,C0、D0、D3、E0和E1的AIC值较小,作为备选遗传模型。适合性检验表明,在30个统计量中,C0、D0、D3和E0均只有2个统计量达到显著水平,而E0的AIC值最小,所以选择E0作为叶面积最优模型,即2对加性-显性-上位性主基因+加性-显性-上位性多基因模型。
2.2.4 叶长遗传模型 根据 AIC准则,C0、D3、D4和E1的AIC值较小,作为备选遗传模型。适合性检验表明,在30个统计量中,E1有11个统计量达到显著水平,C0、D3和D4均有2个统计量达到显著水平,但C0的AIC值最小,所以选择C0作为叶长最优遗传模型,即加性-显性-上位性多基因遗传模型。
2.2.5 叶宽遗传模型 根据AIC准则,B1、C0和E1的AIC值较小,作为备选遗传模型。适合性检验表明,在30个统计量中,C0只有1个统计量达到显著水平,综合确定C0为叶宽最优遗传模型,即加性-显性-上位性多基因模型。
2.2.6 叶形指数遗传模型 根据 AIC准则,C0、D0、D3、E0和E1的AIC值较小,作为备选遗传模型。适合性检验表明,在30个统计量中,C0、D0、E0和 E1均有 2个统计量达到显著水平,而D0的AIC值最小,所以D0为叶形指数最优遗传模型,即 1对加性-显性主基因+加性-显性-上位性多基因模型。
2.2.7 茎围遗传模型 根据 AIC准则,B1、D3、D4、E1和E3的AIC值较小,作为备选遗传模型。适合性检验表明,在30个统计量中,D3和D4均没有统计量达到显著水平,而D3的AIC值最小,所以选择D3作为茎围最优遗传模型,即1对完全显性主基因+加性-显性多基因模型。
2.2.8 节距遗传模型 根据 AIC准则,C0、D0、E0和E1的AIC值较小,作为备选遗传模型。适合性检验表明,在30个统计量中,C0、D0、E0和E1均没有统计量达到显著水平,选择AIC值最小的C0为节距最优遗传模型,即加性-显性-上位性多基因模型。
2.2.9 移栽至中心花开放天数遗传模型 根据AIC准则,B1、E0、E1和E3的AIC值较小,作为备选遗传模型。适合性检验表明,在30个统计量中,E1只有3个统计量达到显著水平,综合考虑,选择E1作为移栽至中心花开放天数最优遗传模型,即2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型。
2.2.10 鲜叶重遗传模型 根据AIC准则,D3、E0和E1的AIC值较小,作为备选遗传模型。适合性检验表明,在30个统计量中,E0模型只有1个统计量达到显著水平,综合确定 E0为鲜叶重最优遗传模型,即 2对加性-显性-上位性主基因+加性-显性-上位性多基因模型。
2.2.11 比叶重遗传模型 根据AIC准则,C0、D0、D3、D4和E5的AIC值较小,作为备选遗传模型。适合性检验表明,在30个统计量中,C0、D0、D3和D4模型均没有统计量达到显著水平,而D3的AIC值最小,综合确定D3为比叶重最优遗传模型,即1对完全显性主基因+加性-显性多基因模型。
2.2.12 蒴果重遗传模型 根据AIC准则,C0、D0和E0的AIC值较小,作为备选遗传模型。适合性检验表明,在30个统计量中,C0、D0和E0均只有1个统计量达到了显著水平,而C0的AIC值最小,综合确定C0为蒴果重最优遗传模型,即加性-显性-上位性多基因模型。
2.3 性状遗传效应预测及遗传参数估计
2.3.1 株高遗传效应分析 丸叶×Coker319组合的株高受2对加性-显性-上位性主基因+加性-显性-上位性多基因控制(表2),第1对主基因加性效应为19.65,显性效应为5.99,第2对主基因加性效应为18.76,显性效应为5.96,第1对主基因和第2对主基因加性×加性上位性效应为-14.44,第 1对主基因和第 2对主基因加性×显性上位性效应为-2.10,第1对主基因和第2对主基因显性×加性上位性效应为-1.26,第1对主基因和第2对主基因显性×显性上位性效应为-27.89,可见株高遗传以加性效应及显性×显性上位性效应为主,但二者方向相反;B1、B2和 F2世代的主基因遗传率分别为41.85%、51.71%和 57.53%;多基因遗传率分别为25.50%、9.73%和16.18%。
2.3.2 叶数遗传效应分析 叶数受 2对加性-显性-上位性主基因+加性-显性-上位性多基因控制(表2),第 1对主基因和第 2对主基因加性效应均为-2.65,显性效应均为-0.95,显性效应只有加性效应的35.85%,加性×加性上位性效应为2.03,加性×显性上位性效应与显性×加性上位性效应均为0.56,显性×显性上位性效应为 3.17,可见叶数遗传以加性效应及显性×显性上位性效应为主,但二者方向相反,表现为正向中亲优势;B1、B2和 F2世代的主基因遗传率分别为 36.91%、51.60%和42.63%;多基因遗传率分别为 31.00%、16.84%和42.08%。
2.3.3 叶面积遗传效应分析 叶面积受 2对加性-显性-上位性主基因+加性-显性-上位性多基因控制(表2),第1对主基因与第2对主基因的加性效应基本相当,均稍大于其显性效应,加性×加性上位性效应与加性×显性上位性效应、显性×加性上位性效应、显性×显性上位性效应也基本相当,其中上位性效应>加性效应>显性效应,上位性效应与加性效应及显性效应方向相反,表现为正向中亲优势;B1、B2和F2世代的主基因遗传率分别为2.13%、50.92%和30.32%;多基因遗传率分别为19.53%、13.26%和12.18%。
2.3.4 叶长遗传效应分析 叶长受加性-显性-上位性多基因控制(表3),B1、B2和F2世代的多基因遗传率分别为38.35%、71.54%和60.75%。
2.3.5 叶宽遗传效应分析 叶宽受加性-显性-上位性多基因控制(表3),B1、B2和F2世代的多基因遗传率分别为46.66%、69.05%和62.14%。
2.3.6 叶形指数遗传效应分析 叶形指数受1对加性-显性主基因+加性-显性-上位性多基因控制(表4),主基因加性效应与显性效应基本相当,表现为正向中亲优势;B1、B2和 F2世代的主基因遗传率分别为0.00%、35.43%和49.64%,多基因遗传率分别为0.00%、46.95%和24.00%。
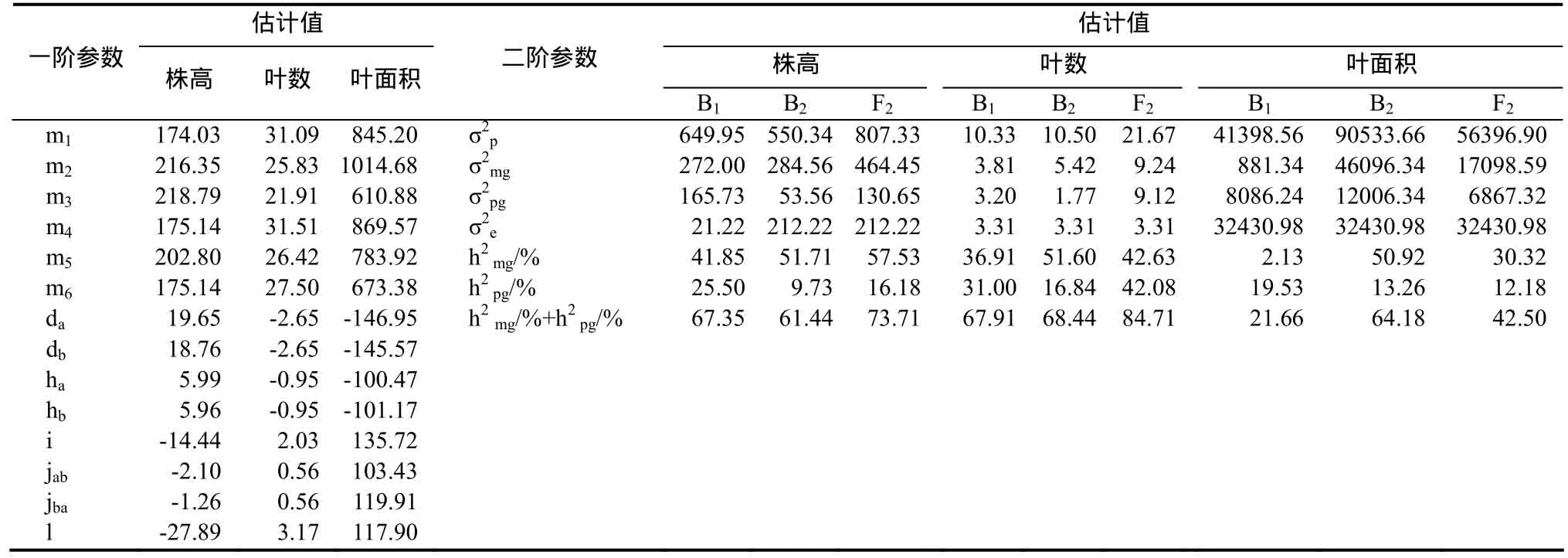
表2 株高、叶数和叶面积的E0模型遗传参数Table 2 The estimate of plant height, leaf number and leaf area in E0 model

表3 叶长、叶宽、节距和蒴果重的C0模型遗传参数Table 3 The estimate of leaf length, leaf width, intermodal distance and capsule weight in C0 model
2.3.7 茎围遗传效应分析 茎围受1对完全显性主基因+加性-显性多基因控制(表4),主基因加性效应和显性效应均为0.02,多基因加性效应为-0.43,多基因显性效应为-0.58,可见茎围遗传以多基因为主,其多基因加性效应和显性效应大小相当;B1、B2和F2世代的主基因遗传率分别为8.84%、0.53%和2.48%;多基因遗传率分别为46.48%、64.12%和53.97%。
2.3.8 节距遗传效应分析 节距受加性-显性-上位性多基因控制(表3),B1、B2和F2世代的主基因遗传率均为0.00%,多基因遗传率分别为65.94%、68.42%和75.08%。
2.3.9 移栽至中心花开放天数遗传效应分析 移栽至中心花开放天数受 2对加性-显性-上位性主基因+加性-显性多基因控制(表5),第1对主基因加性效应为-7.90,显性效应为-2.66,显性效应只占加性效应的33.67%,第2对主基因加性效应为6.69,显性效应为0.25,显性效应只占加性效应的3.74%,第1对主基因与第2对主基因方向相反,加性×加性上位性效应为-8.39,加性×显性上位性效应为0.53,显性×加性上位性效应为-0.90,显性×显性上位性效应为-6.34,多基因加性效应为-2.80,多基因显性效应为-1.44,可见移栽至中心花开放天数遗传以加性×加性上位性效应、加性效应及显性×显性上位性效应为主,亲本型上位性效应大于重组型上位性效应,主基因和多基因均表现为正向中亲优势;B1、B2和F2世代的主基因遗传率分别为50.01%、59.73%和 64.79%;多基因遗传率分别为 0.00%、7.24%和9.87%。
2.3.10 鲜叶重遗传效应分析 鲜叶重受2对加性-显性-上位性主基因+加性-显性-上位性多基因控制(表5),第1对主基因和第2对主基因加性效应均为-7.90,显性效应均为-7.79,加性×加性上位性效应为7.47,加性×显性上位性效应为7.54,显性×加性上位性效应为7.56,显性×显性上位性效应为9.51,可见鲜叶重遗传几种效应大致相当,其中上位性效应>加性效应>显性效应,上位性效应与加性效应及显性效应方向相反,表现为正向中亲优势;B1、B2和 F2世代的主基因遗传率分别为 1.36%、55.38%和44.26%;多基因遗传率分别为31.38%、18.45%和3.18%。
2.3.11 比叶重遗传效应分析 比叶重受 1对完全显性主基因+加性-显性多基因控制(表4),主基因加性效应和显性效应均为-0.0025,多基因加性效应为-0.0022,多基因显性效应为-0.0019,可见比叶重遗传主基因和多基因的加性效应和显性效应大致相当;B1、B2和 F2世代的主基因遗传率分别为0.60%、1.05%和 38.71%;多基因遗传率分别为62.66%、77.66%和25.36%。
2.3.12 蒴果重遗传效应分析 蒴果重受加性-显性-上位性多基因控制(表3),B1、B2和F2世代的多基因遗传率分别为56.27%、59.04%和82.34%。
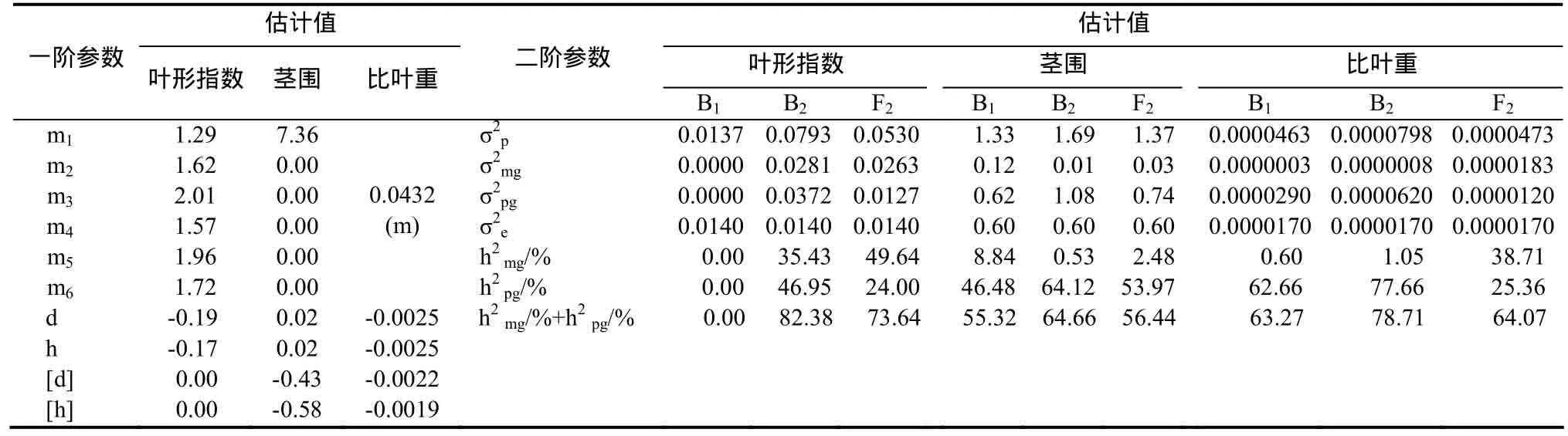
表4 叶形指数的D0模型和茎围、比叶重的D3模型遗传参数Table 4 The estimate of genetic parameters of leaf index in D0 model and stem girth, leaf mass per area in D3 model

表5 移栽至中心花开放天数的E1模型和鲜叶重的E0模型遗传参数Table 5 The estimate of genetic parameters of days of transplanting to flowering in E1 model and fresh leaf weight in E0 model
3 讨 论
3.1 性状遗传率比较
在F2世代中,按照遗传率大小各植物学性状排序依次为:叶数、蒴果重、节距、移栽至中心花开放天数、株高、叶形指数、比叶重、叶宽、叶长、茎围、鲜叶重和叶面积,几个相关性状的遗传率大小、顺序与牛佩兰[8-10]和艾树理[11]的研究结果基本一致,其中株高结果与王日新[12]的研究结果接近。按照主基因遗传率大小各性状排序依次为:移栽至中心花开放天数、株高、叶形指数、鲜叶重、叶数、比叶重、叶面积和茎围,而叶长、叶宽、节距和蒴果重没有检测到主基因的存在。移栽至中心花开放天数、株高、叶形指数、鲜叶重、叶数、比叶重及叶面积的主基因遗传率较高,适合在早代选择,而茎围、叶长、叶宽、节距和蒴果重则宜在晚代选择。
3.2 性状基因效应分析
株高遗传以加性效应及显性×显性上位性效应为主,这与肖炳光[13]、卢秀萍[15]的研究结果一致,后者与王日新[12]及Matzinger[2]的研究结果一致;叶数遗传以加性效应及显性×显性上位性效应为主,其中前者与Matzinger[2]、艾树理[11]、巫升鑫[5]和陈顺辉[6]的研究结果一致,后者与牛佩兰[10]的研究结果不一致,为首次发现;叶面积几种遗传效应差不多,其中上位性效应>加性效应>显性效应;叶形指数主基因加性效应与显性效应基本相当;茎围遗传以多基因为主,其多基因加性效应和显性效应大小相当,这与许健[14]和牛佩兰[10]研究结果不一致;移栽至中心花开放天数遗传以加性×加性上位性效应、加性效应及显性×显性上位性效应为主,其中前者与大多数研究者的结果不一致,中者与Matzinger[2]的研究结果一致,后者与Matzinger[2]和Pandeya[3]的研究结果不一致;鲜叶重几种遗传效应大致相当,其中上位性效应>加性效应>显性效应;比叶重遗传主基因和多基因的加性效应和显性效应大致相当;而叶长、叶宽、节距和蒴果重没有检测到主基因的存在。
本研究还发现,上位性效应是一种非常重要的遗传效应,如在株高、叶数、叶面积、移栽至中心花开放天数及鲜叶重的杂种优势形成过程中起到非常重要的作用,一些作用方向相同,而另一些作用则方向相反,说明基因聚合后会产生不同的互作效应。另外在常规选育中,一般认为只有加性效应可以稳定遗传,选择育种中得到的可稳定遗传的变异就是加性效应值。事实上,对于烟草来说,加性×加性上位性效应可以通过选择稳定遗传,如果加性×加性上位效应存在,那么在选择育种中将会产生额外的遗传收益。在移栽至中心花开放天数中,加性×加性上位效应占据主导地位,可以通过选择稳定地遗传杂种优势。通过上位性效应分析,还可以为育种亲本的选择提供更为详细的信息。即通过对选育亲本的上位性遗传效应预测,选择既有较大加性效应,又有较高上位性效应的育种亲本,以期获得更理想的育种效果。
本研究结果与前人研究有相同也有不同,前人研究基本不能像孟德尔方法那样追踪个别基因,而只能从一组基因的总体上得到有关遗传效应及其相对重要的信息。对植物遗传改良来说,十分需要有关数量性状遗传体系以及其中个别重要基因的信息,以便通过育种手段进行遗传操作。“主基因+多基因”混合遗传模型的出现,有效地解决了上述问题。
4 结 论
烤烟的株高、叶数、叶面积和鲜叶重均受2对加性-显性-上位性主基因+加性-显性-上位性多基因(E0)控制,移栽至中心花开放天数受两对加性-显性-上位性主基因+加性-显性多基因(E1)控制,茎围和比叶重均受1对完全显性主基因+加性-显性多基因(D3)控制,叶形指数受 1对加性-显性主基因+加性-显性-上位性多基因(D0)控制,叶长、叶宽、节距和蒴果重均受加性-显性-上位性多基因(C0)控制,移栽至中心花开放天数、株高、叶形指数、鲜叶重、叶数、比叶重及叶面积的主基因遗传率较高,适合在早代选择,而茎围、叶长、叶宽、节距和蒴果重则宜在晚代选择。
[1]Gopinath D M, Ramanarao V V, Subrahmanyam M, et al.A study of diallel crosses between varieties of Nicotiana tabacum L.for yield component[J].Euphytica, 1966,15(2): 171-178.
[2]Matzinger D F, Mann T J, Cocherham C C.Diallel crosses in Nicotiana tabacum[J].Crop Science, 1962,2(2): 383-386.
[3]Pandeya R S, Dirks V A, Poushinsky G.Quantitative genetic studies in flue-cured (Nicotiana tabacum).Ⅰ.Agronomic characters[J].Canadian Journal of Genetics and Cytology, 1983, 25(2): 336-345.
[4]Wiklinson C A, Tilson W M.Diallel analysis of crosses among Virginia fire-cured tobacco cultivars[J].Tobacco Science, 1994, 38(1): 21-24.
[5]巫升鑫,潘建菁,陈顺辉,等.烤烟若干农艺性状的杂种优势及其遗传分析[J].中国烟草学报,2001,7(4):17-22.
[6]陈顺辉,巫升鑫,倪金应,等.烤烟主要数量性状的配合力研究[J].中国烟草学报,2004,10(3):25-28.
[7]王志德,王元英,牟建民.烟草种质资源描述规范和数据标准[M].北京:中国农业出版社,2006.
[8]牛佩兰,佟道儒,骆启章.烤烟几个主要数量性状遗传力的估算[J].中国烟草,1980(1):13-14.
[9]牛佩兰,刘洪祥,刘伟.烤烟几个主要数量性状相关遗传力的初步研究[J].中国烟草,1984(4):4-5.
[10]牛佩兰,佟道儒.烟草几个主要农艺性状的基因效应分析[J].中国烟草,1989(1):9-12.
[11]艾树理,王秀蓉.烤烟数量性状配合力和遗传力研究[J].烟草科技,1984(3):40-46.
[12]王日新,任民,张兴伟,等.普通烟草栽培种内株高性状主基因加多基因遗传分析[J].中国烟草科学,2009,30(2):15-20.
[13]肖炳光,朱军,卢秀萍,等.烤烟主要农艺性状的遗传与相关分析[J].遗传,2006,28(3):317-323.
[14]许健,杨德,张锦伟.烤烟亲本配合力的双列杂交分析[J].烟草科技,2004(l):29-32.
[15]卢秀萍,肖炳光.烤烟株高的发育遗传研究[J].中国烟草学报,2006,12(4):31-34.
猜你喜欢
农业工程学报(2022年13期)2022-10-09
计算机技术与发展(2021年2期)2021-03-08
临床肝胆病杂志(2020年1期)2020-12-20
聊城大学学报(自然科学版)(2020年1期)2020-11-27
科学导报(2020年51期)2020-09-09
消费导刊(2017年24期)2018-01-31
中国科技纵横(2016年15期)2016-12-29
甘肃教育(2016年22期)2016-12-20
中国茶叶加工(2015年3期)2015-02-27
中国茶叶加工(2015年3期)2015-02-27
