
NLRP3炎性小体在EAE大鼠中的表达及意义
2012-01-26 02:56银晓端周建波
中风与神经疾病杂志 2012年8期
银晓端, 曾 丽, 周建波
多发性硬化(multiple sclerosis,MS)是一种累及中枢神经系统白质并且以炎性脱髓鞘为特征的自身免疫性疾病,本病病因至今尚未完全清楚,炎症通常被认为在自身免疫性疾病的发病过程中起了至关重要的作用。因此,对炎症的相关研究是探索MS发病机制中的重要方面。近年来发现,许多感染性疾病和自身免疫疾病的发生与NLRP3炎性小体介导的炎症反应关系密切[1],但目前为止,国内尚未见NLRP3炎性小体与MS或EAE关系的研究报道。本实验拟从NLRP3炎性小体入手,通过对MS的经典动物模型实验性自身免疫性脊髓炎大鼠(EAE)的研究,探索其是否参与MS炎性反应的发生,以及其上下游参与炎性反应发生的靶分子caspase-1、IL-1β细胞因子的表达情况。阐明NLRP3炎性小体信号通路在MS炎症反应中的作用机制,将可能为MS的诊断和治疗提供新的思路。
1 材料和方法
1.1 实验动物和分组 选取10只健康豚鼠,体重300~400g,雌雄不限,用于制备抗原。健康纯种远交雌性Wistar大鼠30只,体重180~220g,8~10周龄。将大鼠随机分为两组,分别为正常对照组10只、EAE模型组20只。以上动物均由广西医科大学动物中心提供。
1.2 EAE模型制备 用10%水合氯醛按0.5 ml/100g将豚鼠麻醉后断头处死,在冰上迅速解剖分离出脊髓,并去除脊髓表面的脊膜,再称重,加入等量4℃的0.9%生理盐水,匀浆后制成50%的脊髓匀浆液(GPSCH),再加入等量的含6mg/ml卡介苗的完全弗式佐剂(CFA,Sigma公司),混匀后用玻璃注射器抽打制成油包水型乳剂抗原GPSCH-CFA。
1.3 动物免疫和干预 将大鼠用10%水合氯醛按0.5ml/100g麻醉,模型组每只大鼠分别给予0.5ml的GPSCH-CFA,于双后足垫真皮下和颈部皮下3点进行皮下注射;对照组在相同部位注射0.5ml的CFA混合剂(完全弗氏佐剂和生理盐水按1∶1等体积混合,不含GPSCH)。免疫当天计为0d。
1.4 EAE模型成功标准 从0d起每日上午观察大鼠行为学方面的变化,同时测量体重并详细记录。神经功能评分采用7分评分法[2]:0分:不发病;1分:动物皮毛不光整,尾部无力;2分:尾巴瘫痪无力;3分:尾巴瘫痪无力加后肢无力;4分:尾部瘫痪无力加部分后肢瘫痪;5分:后肢完全瘫痪;6分:后肢+前肢瘫痪;7分:濒死或死亡状态。
1.5 组织病理学观察 对照组在免疫后第21天统一处死,模型组评分1分以上的发病大鼠在发病后病情高峰时(大鼠神经症状持续不增而体重增加,或神经症状开始缓解)处死大鼠,用10%水合氯醛按0.5ml/100g进行腹腔注射麻醉,先后用生理盐水及4%多聚甲醛常规灌注,解剖分离出大脑、脑干、脊髓放在4%多聚甲醛中固定24h,然后逐级酒精脱水、包埋、切片,常规HE染色,砂罗硌花青法髓鞘染色,光学显微镜观察病理改变和脱髓鞘情况。
1.6 免疫组化 SP法检测 NLRP3、caspase-1、IL-1β的表达。兔抗大鼠NLRP3、caspase-1、IL-1β多克隆抗体为北京博奥森试剂,SP免疫组化检测系统及DAB显色剂均购自北京中杉金桥有限公司。将各部位标本经石蜡包埋后作5μm连续切片,余下步骤按免疫组化试剂盒说明进行,以细胞膜或胞浆黄色或棕黄色为阳性。阴性对照采用0.01mmol/L的PBS代替一抗,其余方法相同。所有大鼠大脑、脑干、脊髓各取3张切片,每张切片取互不重叠的5个400×倍视野。采用Image-Pro Plus 6.0软件对图像进行分析,计数各视野的阳性细胞数(个/视野)并计算其平均光密度(积分光密度/面积,IOD/area)。
2 结果
2.1 一般情况观察 所有大鼠免疫后的前3d均有体重下降。对照组大鼠3d后体重开始回升并持续增加,无1例发病。模型组免疫后第13天开始发病,伴体重持续下降,各大鼠发病后反应迟钝、毛色晦暗、尾部无力,部分出现肢体瘫痪。模型组发病13只,发病率为65%,无1只死亡。EAE模型组高峰时发病症状的评分为4.56±1.43。
2.2 组织病理学改变 HE染色可见:模型组发病大鼠大脑、脑干、脊髓内有不同程度的淋巴细胞浸润,部分小血管呈袖套样改变。对照组未见明显炎性细胞浸润(见图1)。髓鞘染色可见:模型组发病的大鼠脑组织和脊髓结构疏松,呈弥漫性脱髓鞘改变,而对照组大鼠未见异常改变(见图1)。
2.3 NLRP3的表达 各部位阳性细胞数比较,大脑、中脑、脊髓模型组均高于正常组(P<0.05)。各部位平均光密度值相比,中脑和脊髓模型组高于对照组(P<0.05)(见表1、图2)。
2.4 caspase-1的表达 各部位细胞数比较,大脑、中脑、脊髓模型组均高于正常组(P<0.05)。平均光密度值相比,各部位平均光密度值相比,中脑和脊髓模型组高于对照组(P<0.05)(见表2、图2)。
2.5 IL-1β的表达 各部位细胞数比较,大脑、中脑、脊髓模型组均高于正常组(P<0.05)。各部位平均光密度值相比,大脑、中脑和脊髓模型组高于对照组(P<0.05)(见表3、图2)。
2.6 NLRP3、caspase-1、IL-1β 的相关性检测分别对正常组、模型组的各项指标间的平均光密度值进行相关性检验。正常组的各指标之间的表达量均无相关性。而模型组的NLRP3与caspase-1的表达有明显相关(r=0.715,P<0.05),而与 IL-1β 的表达无明显相关(P>0.05);同时模型组的caspase-1与IL-1β 的表达也明显相关(r=0.821,P<0.05)。
表1 NLRP3的表达(±s)
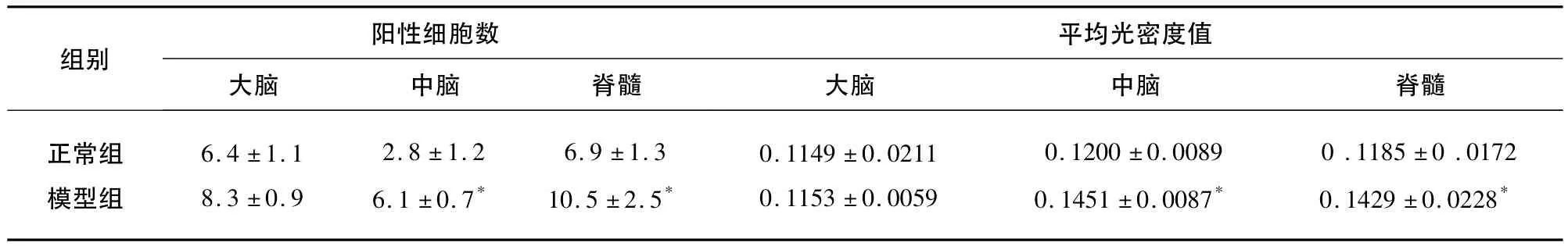
表1 NLRP3的表达(±s)
与正常组比较*P<0.05
阳性细胞数平均光密度值组别大脑 中脑 脊髓正常组模型组大脑 中脑 脊髓6.4 ±1.1 8.3 ±0.9 2.8 ±1.2 6.1 ±0.7*6.9 ±1.3 10.5 ±2.5*0.1149 ±0.0211 0.1153 ±0.0059 0.1200 ±0.0089 0.1451 ±0.0087*0.1185 ±0.0172 0.1429 ±0.0228*
表2 caspase-1的表达(±s)
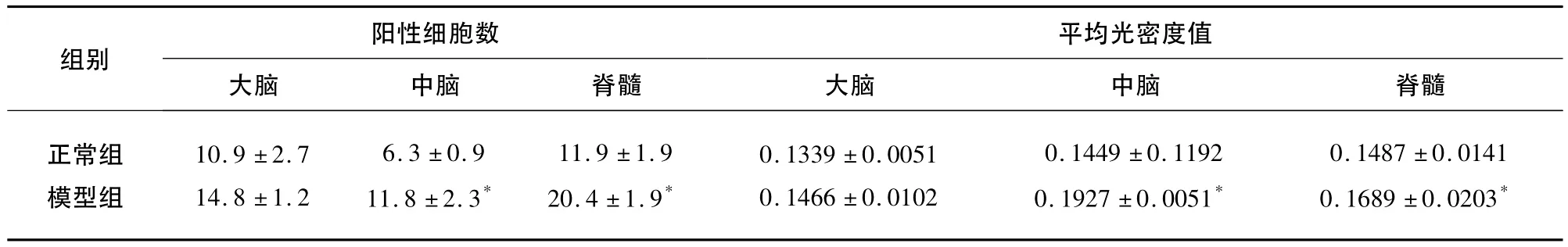
表2 caspase-1的表达(±s)
与正常组比较*P<0.05
阳性细胞数平均光密度值组别大脑 中脑 脊髓正常组模型组大脑 中脑 脊髓10.9 ±2.7 14.8 ±1.2 6.3 ±0.9 11.8 ±2.3*11.9 ±1.9 20.4 ±1.9*0.1339 ±0.0051 0.1466 ±0.0102 0.1449 ±0.1192 0.1927 ±0.0051*0.1487 ±0.0141 0.1689 ±0.0203*
表3 IL-1β 的表达(±s)
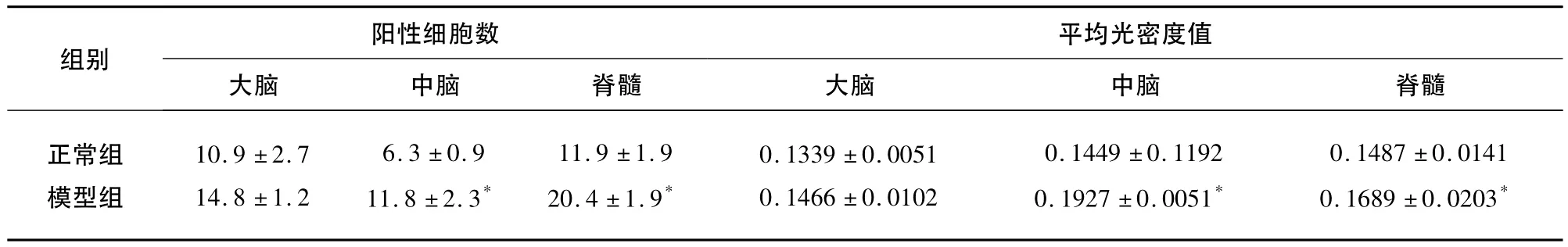
表3 IL-1β 的表达(±s)
与正常组比较*P<0.05
阳性细胞数平均光密度值组别大脑 中脑 脊髓正常组模型组大脑 中脑 脊髓10.9 ±2.7 14.8 ±1.2 6.3 ±0.9 11.8 ±2.3*11.9 ±1.9 20.4 ±1.9*0.1339 ±0.0051 0.1466 ±0.0102 0.1449 ±0.1192 0.1927 ±0.0051*0.1487 ±0.0141 0.1689 ±0.0203*
3 讨论
NLRPs蛋白家族在调节天然免疫和获得性免疫方面具有重要意义,其通过识别病原相关分子模式能迅速启动先天性免疫,在机体的免疫防御中发挥重要作用,其在先天性免疫中参与炎症反应和细胞凋亡,还与一些自身免疫性疾病的发生密切相关[3]。NLRPs蛋白家族中已经报道通过形成炎性小体发挥功能的分子主要有4个,包括NLRP1、NLRP3、IPAF和AIM-2。NLRP3炎性小体是目前研究最多的一种炎性小体,其作为天然固有免疫的重要组分,能被多种类型的病原体或危险信号所激活,因此NLRP3炎性小体在多种疾病过程中都发挥了关键作用,从最初被确认的家族性周期性自身炎症反应到2型糖尿病、阿尔海默茨病和动脉粥样硬化症等[4]。
国外有研究发现NLRP3基因敲除后EAE的潜伏期延长、临床症状改善、髓鞘脱失减轻、炎性细胞浸润减少、星形胶质细胞增生减少[5]。Guarda等的研究还发现,I型干扰素用于治疗MS的作用机制之一,就是其能通过减少NLRP3等炎性小体的激活来减少IL-l的生成[6]。由此可见,NLRP3可能参与MS的发病,但是NLRP3在EAE上的作用未见有相关研究验证。此外,NLRP3已在很多疾病研究中被证实会表达增高,至于在EAE中是否表达升高,其意义又如何,国内未见有相关报道,因此,本实验对该方面进行了相关探索。
在本次实验中,我们用豚鼠脊髓匀浆作为抗原免疫Wistar大鼠成功诱导了EAE做为MS模型,该MS模型发病病程呈多样性,而且炎症反应较重[7],因此适合用做研究EAE炎性反应的研究对象。本次实验中对发病大鼠的症状观察、HE染色和髓鞘染色均发现,该模型的临床症状、炎性细胞浸润、髓鞘脱失情况均符合MS的病变特点。同时免疫组化结果表明,模型组的 NLRP3、caspase-1、IL-lβ 与对照组对比有显著差异,提示 NLRP3、caspase-1、IL-lβ 与EAE的发病过程、炎症浸润和脱髓鞘关系密切,这也提示了 NLRP3、caspase-1、IL-lβ 可能参与了 MS发病。NLRP3在EAE中表达上调目前认为有两种可能:(1)NLRP3作为Nod样受体亚家族成员之一主要表达在单核巨噬细胞、T细胞、B细胞、粒细胞、树突状细胞等炎性细胞中[8]。NLRP3在EAE的脑组织和脊髓的表达随着大量炎性细胞的浸润而增多。(2)炎症中一些危险信号和代谢产物如细胞坏死产物、异常代谢产物等,以及EAE制模过程中使用的免疫佐剂的卡介苗,都能不同程度地激活NLRP3[9],从而导致其表达增多。
本次实验还发现,caspase-1、IL-lβ在EAE的表达增多,且 EAE的 NLRP3与 caspase-1、IL-1β与caspase-1的表达量均有相关性,与NLRP3炎性小体的作用机制有关:NLRP3被激活后会形成蛋白复合物-NLRP3炎性小体,同时活化 caspase-1,活化的caspase-1再对pro-IL-1β、pro-IL-18等细胞因子底物进行剪接,促进IL-lβ、IL-18等生成和分泌[10]。但是不仅是NLRP3炎性体,还有诸如NLRP1等其他炎性体能激活caspase-1。因此本实验中模型组的IL-1β与NLRP3相关性无统计学意义,可能是EAE发病中还有除了NLRP3之外的其它炎性体被激活,导致后来IL-lβ的形成与NLRP3表达量的相关性不明显。被caspase-1处理后生成的IL-lβ、IL-18同属于IL-l超家族,二者都是重要的促炎性因子,都能通过结合相应的受体激活下游的炎性通路,从而促进如IL-3、IL-5、IL-6、IL-13等炎症因子的释放。但是它们之间还是有所区别的:IL-lβ能促进 T细胞的增殖,在Th17细胞分化中起关键作用[11];IL-18能够诱导IFN-γ的表达和分泌,刺激Th1或Th2细胞介导的免疫反应[12]。上述观点与Gris等的研究结论一致,他们发现缺乏NLRP3会导致EAE的IFN-γ和IL-17合成和分泌的减少,提示NLRP3炎性小体能通过增加IL-lβ和IL-18来刺激Th17细胞和Th1细胞的增殖和分化,参与EAE的炎症反应[5]。
本研究为NLRP3炎性体和IL-1β在EAE大鼠的发病中起到重要作用这一假说提供了有力证据。但MS炎性反应的发生机制复杂,目前普遍认为是细胞免疫介导的自身免疫疾病,同时本次实验结果还提示可能还有其他炎性小体参与了EAE的发病,因此还应研究NLRP3与其他炎性小体的之间的相互联系,并观察NLRP3对各亚型淋巴细胞变化的影响,探讨NLRP3在EAE中的变化规律和作用机制,以便为研究MS的研究提供新的切入点,完善MS的发病机制。
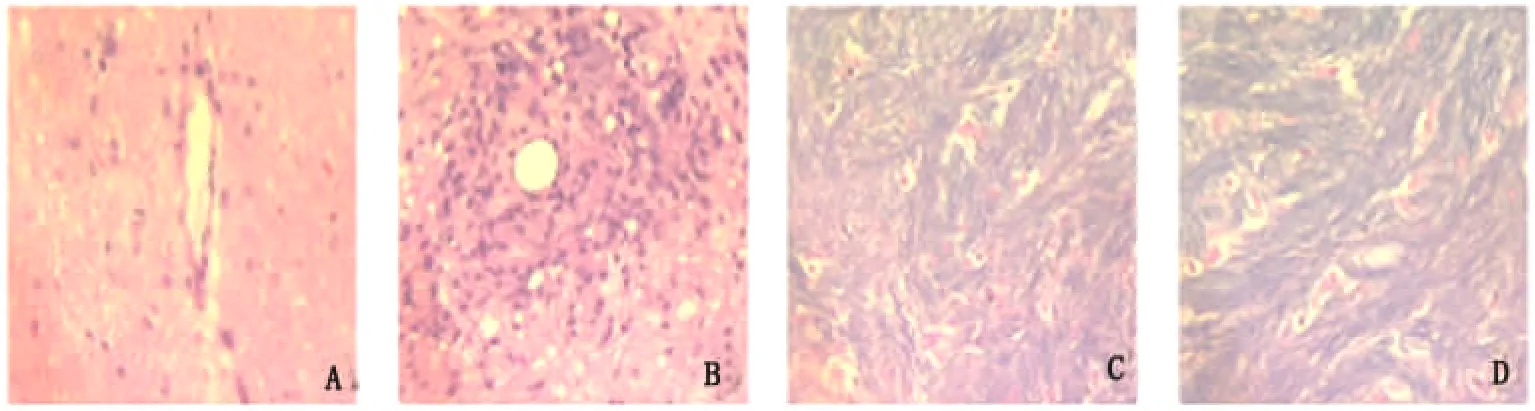
图1 A:正常组HE;B:模型组HE;C:正常组髓鞘;D:模型组髓鞘(病理染色×400)
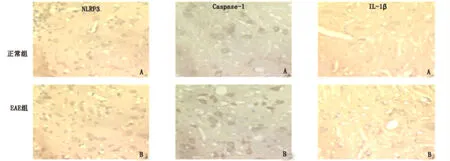
图2 A:正常组;B:模型组(从左到右依次为NLRP3、caspase-1、IL-1β免疫组化染色×400)
[1]Strowig T,Henao-Mejia J,Elinav E,et al.Inflammasomes in health and disease[J].Nature,2012,481(7381):278 -286.
[2]Hooper DC,Spitsin S,Kean RB,et al.Uric acid,a natural scavenger of peroxynitrite,in experimental allergic encephalomyelitis and multiple sclerosis[J].Proc Natl Acad Sci USA,1998,95(2):675 -680.
[3]林福全.固有免疫模式受体NALPs的信号通路及其与自身免疫性疾病的研究进展[J].国际免疫学杂志,2008,31(6):419 -422.
[4]毛开睿,孙 兵.NLRP3炎症小体研究进展[J].现代免疫学,2011,31(1):1 -4.
[5]Gris D,Ye Z,Iocca HA,et al.NLRP3 plays a critical role in the development of experimental autoimmune encephalomyelitis by mediating Th1 and Th17 responses[J].J Immunol,2010,185(2):974 -981.
[6]Guarda G,Braun M,Staehli F,et al.Type I interferon inhibits interleukin-1 production and inflammasome activation[J].Immunity,2011,34(2):213 -223.
[7]Dong M,Liu R,Guo L,et al.Pathological findings in rats with experimental allergic encephalomyelitis[J].APMIS,2008,116(11):972 -984.
[8]Ting JP,Davis BK.CATERPILLER:a novel gene family important in immunity,cell death,and diseases[J].Annu Rev Immunol,2005,23:387-414.
[9]Lalor SJ,Dungan LS,Sutton CE,et al.Caspase-1-processed cytokines IL-1beta and IL-18 promote IL-17 production by gammadelta and CD4 T cells that mediate autoimmunity[J].J Immunol,2011,186(10):5738-5748.
[10]Martinon F,Burns K,Tschopp J.The inflammasome:a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta[J].Mol Cell,2002,10(2):417 -426.
[11]Chung Y,Chang SH,Martinez GJ,et al.Critical regulation of early Th17 cell differentiation by interleukin-1 signaling[J].Immunity,2009,30(4):576 -587.
[12]Sims JE,Smith DE.The IL-1 family:regulators of immunity[J].Nat Rev Immunol,2010,10(2):89 -102.
猜你喜欢
中西医结合心脑血管病杂志(2022年19期)2022-11-19
中国临床解剖学杂志(2022年1期)2022-11-15
检验医学与临床(2022年12期)2022-06-27
中国人兽共患病学报(2022年5期)2022-06-08
哈尔滨理工大学学报(2022年1期)2022-05-10
国际眼科杂志(2022年2期)2022-02-18
中国医学影像技术(2022年1期)2022-02-15
青岛大学学报(医学版)(2021年2期)2021-07-08
中国实用神经疾病杂志(2020年21期)2020-12-04
中国医药导报(2020年12期)2020-05-25
