
缺氧后胚鼠神经元自噬及凋亡相关蛋白的表达
2012-01-26 02:55陈艾,周晖,童煜,毛萌
中风与神经疾病杂志 2012年8期
陈 艾, 周 晖, 童 煜, 毛 萌
缺血缺氧性脑损伤是儿科常见疾病之一,可累及神经系统发育中的多个区域而遗留永久性的损伤,成活者常遗留脑瘫、智力低下等严重后遗症[1,2]。但缺血缺氧性脑损伤的发病机制尚不明确,其有效干预措施也有待探索。自噬是继坏死、凋亡后发现的第3种细胞死亡形式,被称为Ⅱ型程序化死亡。自噬、凋亡、坏死3种现象在细胞应激状态下的发生秩序一直是国内外争论的热点。
本研究旨在观察缺氧致胚鼠神经元损伤后,自噬以及凋亡的发生及变化情况。为进一步研究自噬和凋亡的关系,以及两者对缺血缺氧性脑病的影响提供新的思路。
1 材料与方法
1.1 实验动物 3只17~19d SD孕鼠,清洁级,体质量350~400g,每次使用8只胚鼠,分3次培养神经元。由四川省医学科学院实验动物研究所提供[生产许可证号:SCXK(川)2004-15]。
1.2 主要试剂 多聚-D-赖氨酸、鼠尾胶原(Sigma,美国);高糖培养基、神经A培养基、B-27神经刺激因子、兔源LC3 I/II单克隆抗体、caspase-3单克隆抗体(赛信通,美国);鼠源actin单克隆抗体、二喹啉甲酸(bicinchoninic acid,BCA)蛋白定量试剂盒(中杉金桥,北京);双抗(100IU/ml青霉素、100μg/ml链霉素)、高糖DMEM、10%胎牛血清(Hyclone,美国)。
1.3 主要设备 24孔塑料培养板(洁特生物,广州),二氧化碳(CO2)培养箱(Sanyo,日本),厌氧培养箱(Thermo,美国),超净工作台(Artech,加拿大)。
1.4 原代神经元培养 无菌条件下分离胚鼠大脑皮质,经去除脑膜、胰酶消化后,以1.6~1.8×106/ml的密度接种,接种培养基为高糖DMEM,加10%胎牛血清、1%glutamax、双抗,放入 37℃、5%CO2饱和湿度培养箱中。4h后更换为神经元培养基,之后每3d半量换液,至7d神经元成熟。
1.5 神经元缺氧模型的建立 选用培养7d生长良好的神经元,放入国产厌氧培养箱中培养1~6h,厌氧培养条件为 37℃、1%O2。
1.6 蛋白样品的制备及Western blot检测 提取细胞总蛋白后用BCA法定量。分别取25μg样品进行电泳(15%SDS-PAGE)和转膜(PVDF膜)。先后用 LC3 I/II(1∶1000)、caspase-3(1∶1000)以及Actin(1∶1000)的一抗和辣根过氧化酶标记的二抗(1∶1000)进行杂交,洗膜后使用Immobilon Western化学发光底物(Millipore,美国)在化学发光成像仪(Bio-Rad公司)中显影。所得结果用Gel-pro Analyzer软件分析。
1.7 神经元免疫荧光 选取体外培养7d生长良好的大鼠皮层神经元,吸净培养基,用预热的PBS液洗3次,4%多聚甲醛室温固定15min,PBS洗涤后用 0.2%TritonX-100/PBS 于 4℃ 下处理 15min,5%BSA封闭30min,滴加一抗小鼠源MAP2(1∶200)、GFAP(1∶200),湿盒中4℃孵育过夜,PBS洗涤后滴加抗小鼠的IgG-FITC(1∶200),避光室温孵育1h,DAPI(1∶500)避光1min,PBS充分洗涤后50%缓冲甘油封片,荧光显微镜下观察、拍照。以MAP2阳性细胞占细胞总数的百分比表示神经元细胞纯度。
1.8 统计学分析 数据分析采用SPSS13.0软件,计量资料用均数±标准差(±s)表示,组间比较采用方差分析。检验水准α=0.05,以P<0.05认为差异有统计学意义。
2 结果
2.1 皮质神经元的原代培养
2.1.1 神经元形态学观察 正常神经元呈圆形或椭圆形,胞体发亮,周边可见明显光晕,表明细胞活性很好,突触伸展成多极、双极,交织成网状(见图1A)。OD 3h导致神经元突触回缩,网状结构被破坏(见图1B)。OD 6h后神经元碎片化,突触膨胀断裂,网状结构几乎消失(见图1C)。
2.1.2 免疫荧光鉴定 原代神经元鉴定MAP2标记的成熟神经元显示绿色荧光,荧光显微镜下观察神经元形态结构完整,树突及轴突舒展(见图2A)。OD 3h后神经元胞体不再饱满度,突触回缩断裂,网状结构被破坏(图2B)。OD 6h神经元感光度降低,突触膨胀断裂,网状结构几乎消失(见图2C)。GFAP阳性表达的为星形胶质细胞(见图2D),计数十个视野范围内的100个细胞,星形胶质细胞沾染率<5%。
2.2 缺氧能明显诱导自噬现象的发生 缺氧可以明显诱导自噬的发生。与OD 0.5h皮质神经元对比,随缺氧时间延长,LC3I向LC3II的转换明显增多,尤其在OD 4h时出现高峰,LC3-II/Actin(OD)=0.8228 ±0.012,之后逐渐减弱。至 OD 6h,神经元细胞几乎碎裂。OD 0.5h~OD 3h与OD 4h~OD 6h的LC3-II/actin均值比较有统计学差异(P<0.05)。
缺氧明显诱导自噬发生的同时,凋亡现象也出现。casapase-3表达趋势在 OD 4h内与 LC3-II一致,但OD 4h~OD 6h时,LC3-II的表达不再增加,出现平台期,而caspase-3的表达继续增强(见图3)。
3 讨论
3.1 自噬 自噬是指细胞在自噬相关基因(autophagy related gene,Atg)的调控下利用溶酶体/噬泡降解自身受损的细胞器和大分子物质的过程[3]。自噬在进化过程中有高度的保守性,是继坏死、凋亡后发现的第3种细胞死亡形式,参与了如神经退行性改变、精神分裂等多种神经精神疾病的发病过程[3]。自噬可清除受损的细胞器,同时降解被包裹的蛋白质、核酸等成分,产生氨基酸和核苷酸进入三羧酸循环,产生小分子和能量(ATP),被细胞再利用。这样就实现了细胞本身的代谢需要和某些细胞器的更新。自噬是一种进化保守的细胞反应,它的这种精细调节对维持细胞的自身稳态有重要作用,所以体内几乎所有细胞中均有基础水平的自噬现象存在[4]。
事实上,自噬是一个动态的过程,包括3个阶段:(1)前自噬泡形成:胞浆中大量膜性结构形成;(2)自噬泡形成:膜性结构包裹受损的细胞器等形成独立的小泡;(3)自噬泡降解:自噬泡的外膜与溶酶体膜融合,形成自噬溶酶体,最终溶酶体酶逐渐消化被包裹的各种小分子物质,产生能量供机体再利用[5]。在神经元遭受缺氧损伤后,自噬发挥着双刃剑的作用。一方面,自噬的过度激活可能介导细胞程序性死亡[6];另一方面,自噬的启动可能有助于维持神经元稳态,减少继发损伤,清除已经被损伤的细胞器而保护神经元。
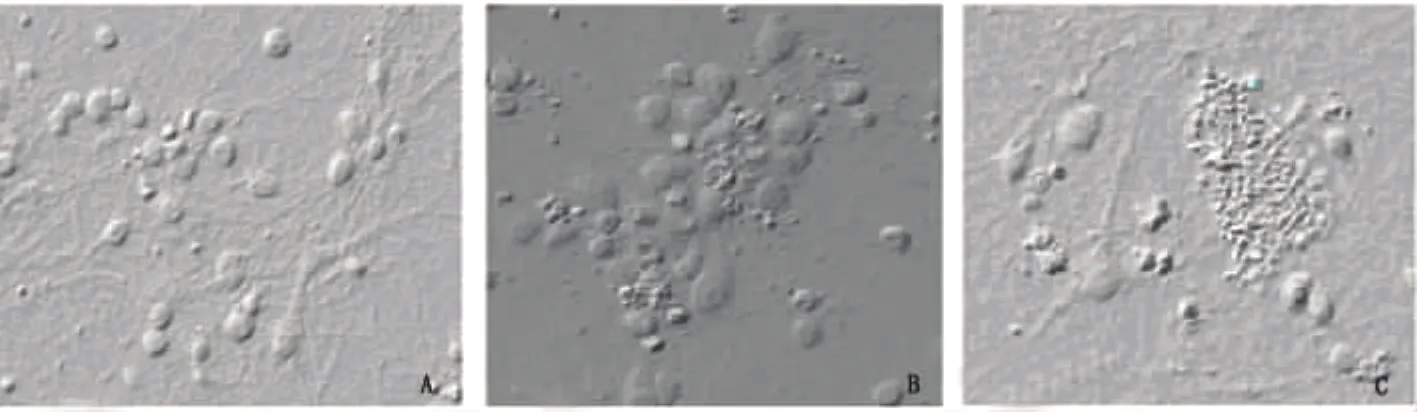
图1 光镜结果(×400)

图2 免疫荧光化学染色(×400)
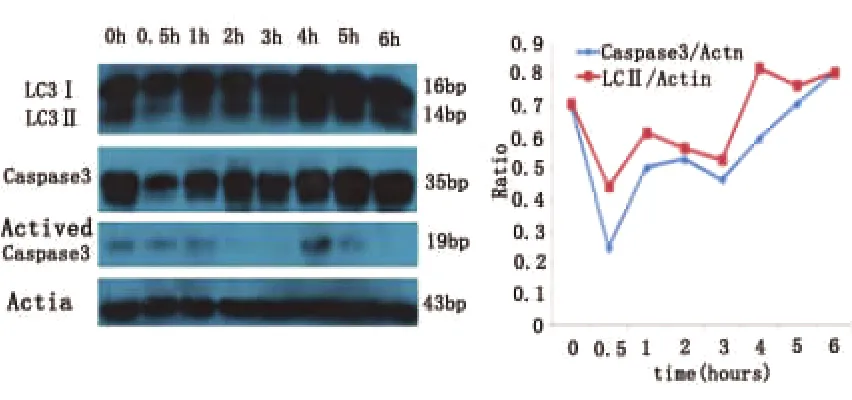
图3 OD处理后皮质神经元LC3II及caspase-3的表达趋势
在所有的Atg蛋白中,目前认为只有LC3/Atg8贯穿于自噬发生的始终,可作为自噬的监测标志[7]。LC3有4种亚型(LC3 I-IV),其中LC3II因表达最多而成为最广泛的自噬监测指标。自噬发生时,LC3的C端被Atg4剪切后链接一个甘氨酸残基即成为LC3I,弥散存在于胞质。再通过类泛素化酶反应,催化偶联磷脂酰乙醇胺(phosphatidylethanolamine,PE),成为 LC3-II(LC3-PE),聚集于自噬体内外膜上。因此,常以LC3II来衡量自噬的发生[8]。
我们已有实验结果证实,脑细胞对缺氧非常敏感。随着缺氧时间的逐步延长,神经元死亡及凋亡的比率明显增加。尤其以缺氧3~5h最为显著,缺氧6h后,神经元几乎死亡[9]。因此,我们选择了神经元缺氧最为敏感的6h内作为观察时间,在神经元遭受缺氧损伤后应用Western blot检测LC3蛋白水平的变化以观察自噬的发生。
我们观察到,未缺氧处理时,LC3II的表达在OD 0h有一个波峰出现,考虑为细胞在提取蛋白的过程中受到了温度、缺氧、细胞破碎等剧烈应激变化所致。
缺氧后的细胞,由于已经有0.5h处于应激耐受状态,可能激发了细胞的稳态,所以 OD 0.5h的LC3II的表达量反而比 OD 0h还低。OD 0.5h~OD4h,LC3II表达相对稳定,处于第一个平台期(P<0.05),OD 0.5h ~OD 4h 组间比较无统计学意义(P>0.05)。随着时间的推移,LC3II表达逐渐增加,于OD 4h跳跃式到达峰值,之后在OD 4h~OD 6h处于第二个平台期,LC3II表达不再上调。OD 4h-OD 6h与OD 0.5h~OD 4h的数值均值比较有统计学意义(P<0.05)。
我们知道,在临床工作中,当卒中、新生儿缺血缺氧性脑病(hypoxic-ischemic encephalopathy,HIE)等缺血缺氧性脑病发生时,最佳的抢救时间是发病后4h以内,被称为治疗的最佳时间窗。之前尚未有研究提示,缺氧应激后4h之内存在自噬现象的发生。以上数据提示,自噬所介导的信号通路,也是缺血缺氧性脑损伤4h内触发的瀑布式病理生理机制之一。此实验结果,佐证了只有尽量争取在缺氧导致起病4h内[1],LC3II处于第一个平台期时,才能得到更好的治疗效果。同时,也提示临床医生,是否能将LC3II的时间依从性表达趋势作为临床判断缺血缺氧性脑损伤预后的依据之一。
3.2 凋亡 caspase-3是凋亡的标志蛋白,一般认为,只有分子量为35bp的caspase-3被激活成17或19bp的小片段时,才能体现凋亡的发生[10]。因此,我们选取被激活的caspase-3(19bp)为检测凋亡的指标。从图3可以看出,缺氧诱导神经元发生自噬的同时,凋亡也被激活,casapase-3(19bp)表达趋势在OD 4h内与LC3II的表达趋势一致。但是,当缺氧应激持续存在,在OD 4h~OD 6h时,LC3-II表达量不再升高,而casapase-3(19bp)表达继续增强。因此,我们认为,当自噬不能维持细胞稳态时,凋亡增强,最终神经元出现坏死。因此,当没能及时在缺氧应激后4h内得到有效治疗的临床患者,病程后4~6h的治疗或许应该以抗细胞凋亡为主。
3.3 小结 目前,对细胞3种程序化死亡现象发生的时间顺序仍有争议[10],有观点认为当细胞遭遇应激时,自噬最先被激活,当自噬不能维持细胞稳态后,细胞再继发凋亡,最后进入坏死,这种顺序是不可逆的[11]。也有观点认为,自噬与凋亡重叠发生,平行执行功能,互为补充,共同维持细胞稳态,抵御应激损伤。本实验研究发现,细胞自噬与凋亡在某段时间内并存,尤其是在应激后的4h内,这为临床干预的“时间窗”学说提供新的理论依据。
本实验研究也提示,既然在缺氧损伤等应激下,细胞会启动自噬机制来维持自身稳态,提高细胞对低氧的耐受力,那么调控自噬这种细胞自身的促生存机制,或许能够作为应对缺氧损伤的新策略。如有文献报道,大鼠局灶性脑缺血再灌注后缺血半暗带脑源性神经营养因子(brain-derivedneurotrophicfactor,BDNF)的mRNA及蛋白表达均明显增加,提示其可能参与了缺血后神经保护[12],那么,提前应用外源性BDNF可能会改善缺氧应激疾病的预后。
其次,监测LC3II的动态变化,可望作为指导临床用药的依据和判断预后的指标。比如,病程4h内治疗以维持细胞稳态,促进自噬为主,预后较好;而当病程进展至4~6h,患者才得以救治的话,治疗应该以抗凋亡为主,且预后相对较差。缺氧启动自噬的信号通路如何,哪些位点参与具体调控,在治疗此类脑损伤的过程中,是否能预防性激活自噬、加强自噬的功能以保护神经元,将是我们实验组下一步研究的内容。
[1]Nelson KB,Lynch JK.Stroke in newborn infants[J].Lancet Neurol,2004,3:150 -158.
[2]Nelson KB.Neonatal encephalopathy:etiology and outcome[J].Dev Med Child Neurol,2005,47:292.
[3]Shpilka T,Elazar Z.Shedding light on mammalian microautophagy[J].Dev Cell,2011,20:1 -2.
[4]Farre JC,Krick R,Subramani S,et al.Turnover of organelles by autophagy in yeast[J].Curr Opin Cell Biol,2009,21:522 -530.
[5]Mehrpour M,Esclatine A,Beau I,et al.Overview of macroautophagy regulation in mammalian cells[J].Cell Res,2010,20:748 -762.
[6]Adhami F,Liao G,Morozov YM,et al.Cerebral ischemia-hypoxia induces intravascular coagulation and autophagy[J].Am J Pathol,2006,169:566 -583.
[7]Mizushima N,Yoshimori T.How to interpret LC3 immunoblotting[J].Autophagy,2007,3:542 -545.
[8]Yang Z,Klionsky DJ.An overview of the molecular mechanism of autophagy[J].Curr Top Microbiol Immunol,2009,335:1 - 32.
[9]周 晖,毛 萌,刘卫平.脑源性神经营养因子对缺氧胚脑皮质神经元的保护作用[J].华西医大学报,2002,33(4):573 -576.
[10]Xue L,Fletcher GC,Tolkovsky AM.Autophagy is activated by apoptotic signalling in sympathetic neurons:analternative mechanism of death execution[J].Mol Cell Neurosci,1999,14:180 - 198.
[11]Edinger AL,Thompson CB.Death by design:apoptosis,necrosis and autophagy[J].Curr Opin Cell Biol,2004,16:663 - 669.
[12]胡跃强,唐 农,刘 泰,等.大鼠局灶性脑缺血再灌注后BDNF mRNA及其蛋白的表达变化[J].中风与神经疾病杂志,2011,28:220-222.
猜你喜欢
大电机技术(2022年3期)2022-08-06
核科学与工程(2021年4期)2022-01-12
智慧医学(2021年2期)2021-09-10
煤气与热力(2021年4期)2021-06-09
保健文汇(2021年5期)2021-05-22
电子产品世界(2021年8期)2021-01-16
中华戏曲(2020年1期)2020-02-12
健康大视野(2019年7期)2019-04-25
现代装饰(2018年5期)2018-05-26
中国社区医师(2017年12期)2017-06-05
