
甘露聚糖结合凝集素诱导DC成熟机制分析①
2012-01-23 12:04王凡平王明永杨建斌赵东方王红坡孙瑞利郭庆合凌明智赵永新宋士军郭继强新乡医学院医学检验系新乡453003
中国免疫学杂志 2012年12期
王凡平 王明永 杨建斌 赵东方 王红坡 邵 峰 孙瑞利 郭庆合 凌明智 赵永新宋士军 郭继强 (新乡医学院医学检验系,新乡453003)
甘露聚糖结合凝集素(Mannan-binding lectin,MBL)是由肝细胞分泌的血浆蛋白,为天然免疫系统中的关键分子[1]。近年来一些研究发现,MBL作为C型凝集素家族的重要成员和天然免疫中的关键分子,除了其固有的抗感染作用外,还发挥多种免疫调节作用[2-7]。陈月等[8]研究发现,MBL 能够诱导树突状细胞(Dendritic cell,DC)成熟,但机制并不明了。已报道,DC成熟过程中,模式识别受体Toll样受体(Toll-like receptors,TLRs)的表达发生改变;文献有报道,与MBL同为胶凝素家族成员的补体C1q能够通过核转录因子NF-κB信号途径诱导DC成熟[9]。MBL诱导DC成熟是否也与TLRs和NF-κB有关呢?需深入研究。
本研究以 TLRs和NF-κB为切入点,旨在初步探讨MBL诱导DC成熟机制。
1 材料与方法
1.1材料 人血浆天然MBL按文献[10]制备;rh-GM-CSF和rhIL-4购自Pepro-Tech公司。PE标记的鼠抗人CD1a、CD80及CD86单克隆抗体(mAb),FITC标记的鼠抗人 CD83、CD86及 MHC-DR的mAb,同型 FITC-IgG1、PE-IgG1阴性参照抗体均为eBioscience公司产品。RPMI1640及尼龙毛柱购自Gibco公司;淋巴细胞分离液(Ficoll,密度1.077)为上海试剂二厂生产;新生牛血清(Newborn calf serum,NCS)购自杭州四季青生物工程材料研究所;鼠抗人 NF-κB mAb p65、抗 β-actin兔多克隆抗体及HRP-羊抗鼠IgG购自Sigma公司;鼠抗人MBL pAb购自R&D公司;核蛋白提取试剂盒为Pierce公司产品,凝胶电泳迁移率变动分析(EMSA)测定试剂盒为Promega公司产品;[γ32-P]ATP为北京亚辉生物试剂公司产品;结合缓冲液:20 mmol/L Tris-HCl(pH8.0)、150 mmol/L NaCl、1 mmol/L CaCl2、2 mmol/L MgCl2、5 g/L BSA、1 g/L 叠氮钠;其它化学试剂均为进口或国产分析纯产品;FACSCalibur流式细胞仪为美国BD公司产品。
1.2方法
1.2.1人外周血Mo的获取 取健康成人志愿者的外周血,肝素抗凝后,用Ficoll密度梯度离心法分离单个核细胞。以PBS洗去血小板,于37℃贴壁3小时后弃上清,以37℃预温的RPMI1640轻洗去除非贴壁的细胞,刮取贴壁的黏附细胞即为Mo,用含10%NCS的 RPMI1640调整细胞密度为1×109个/L。
1.2.2DC的体外诱导 向Mo悬液中加入rhGMCSF和rhIL-4,分3组进行DC培养。常规组:只加rhIL-4和rhGM-CSF培养7天;MBL刺激组:培养5天后加入MBL(10 mg/L)继续培养2天;MBL+Anti-MBL刺激组:培养5天后加入MBL(10 mg/L)和Anti-MBL pAb继续培养2天;人血清白蛋白(HSA)刺激组(阴性对照):培养5天后加入HSA继续培养2天。在诱导DC的过程中,每3天半量换液并补充相应的细胞因子,于倒置显微镜下观察细胞形态。7天后收集悬浮的细胞,分析DC的表型及功能。
1.2.3DC的表型分析 用预冷PBA(0.01 mol/L PBS+20 g/L BSA+0.1 g/L NaN3)洗涤不同实验组诱导7天后的悬浮细胞并调整细胞密度为5×109个/L。于100 μl细胞悬液中,分别加入两两组合的标记抗体,即 PE-抗 CD1a和 FITC-抗 CD83、PE-抗CD80 和 FITC-抗CD40、PE-抗 CD86 mAb 和 FITC-抗MHC-DR mAb各20 μl,同时设 PE和 FITC标记的小鼠IgG1抗体作为对照,于4℃避光标记30分钟后,用预冷PBA洗涤2次。最后用500 μl PBS悬浮细胞,于流式细胞仪上分析DC的表型。
1.2.4制备FITC-MBL 按照文献[11],采用透析法,以FITC标记MBL,标记产物FITC-MBL的F/P比值为2.4。
1.2.5细胞结合试验 imDC用A缓冲液(调整Ca2+浓度分别为 1 mmol/L、5 mmol/L,或者用 5 mmol/L EDTA代替CaCl2)重悬并调整细胞密度为5 ×109个/L,于200 μl细胞悬液中加入 FITC-MBL,37℃避光反应30分钟。选择用人血浆MBL生理浓度的上限15 mg/L作为实验中MBL用量的标准浓度。

表1 引物序列及RT-PCR产物的大小Tab.1 Primer sequence and sizes of RT-PCR products
1.2.6RT-PCR分析DC TLR2、TLR4 mRNA的表达 细胞总RNA的制备:分别取上述体外诱导的各实验组1×107个细胞,用无菌PBS洗1次,按Trizol法操作指南提取总RNA。以1%琼脂糖凝胶电泳鉴定其完整性,用紫外分光光度仪测定其 OD260nm/OD280nm比值。统一调整总RNA浓度为0.15 mg/ml,立即反转录合成cDNA。PCR引物的设计与合成:参考文献[12]并经Primer premier 5.0软件分析,由上海博亚公司合成TLR2、TLR4和GAPDH等3对特异性引物(序列见表1)。GAPDH作为内参照。cDNA合成:分别取不同实验组DC总RNA各1 μg、5 × RT Buffer 4 μl、dNTP Mixture(各 10 mol/L)2 μl、RNase inhibitor(10 U/ μl)1 μl、Oligo(dT)20(10 pmol/ μl)1 μl和 ReverTra Ace 1 μl,加 RNase Free H2O至总反应体积20 μl。42℃反应20分钟合成cDNA第一链,99℃ 5分钟灭活反转录酶终止反应,4℃ 5分钟复性。PCR:反应体系为:10×buffer 2 μl、dNTPs 2 μl(各 0.25 mol/L),上、下游引物各 1 μl(10 μmol/L),模板 cDNA 1.5 μl,Taq DNA 多聚酶(5 U/μl)0.2 μl,加 DEPC 处理水至总反应体积20 μl。TLR2的PCR反应条件为:94℃变性30秒,60℃退火15秒,72℃延伸90秒,循环35次。TLR4的PCR反应条件为:94℃变性30秒,60℃退火25秒,72℃延伸1分钟,循环35次。GAPDH的 PCR反应条件为:94℃变性30秒,63℃退火30秒,72℃延伸1分钟,循环35次。取各PCR扩增产物5 μl于1%琼脂糖凝胶电泳,溴化乙锭染色,紫外灯下观察、拍照,并用凝胶成像仪分析DNA片段的灰度值。
1.2.7EMSA和 Western blot分析 NF-κB细胞核移位 ①EMSA分析:按照Pierce公司提供的试剂盒说明书进行细胞核蛋白的提取,采用Bradford法检测提取液蛋白浓度,核蛋白-70℃冻存备用。根据Promega公司提供的EMSA检测试剂盒说明书进行各组细胞NF-κB活性的检测。探针的制备:分别按顺序取1.75 mol/ml NF-κB特异性核苷酸2 μl、10×T4多聚核苷酸激酶缓冲液1 μl、3 000 Ci/mmol[γ32-P]ATP 1 μl、无核酸酶蒸馏水 5 μl和 5 ×103~1 ×104U/μl的 T4 多聚核苷酸激酶 1 μl,混匀后于37℃水浴中反应10分钟,加入0.5 mol/L EDTA 1 μl终止反应,并加入 89 μl TE 缓冲液(10 mmol/L Tris-HCl,1 mmol/L EDTA,pH7.9),混匀置4℃保存。特异及非特异性竞争实验:在加入标记探针前,先加未标记的特异性探针(NF-κB寡核苷酸探针),并逐渐增加未标记的探针浓度(特异性竞争实验);在加入标记探针前,先加入未标记的非特异性探针,并逐渐增加未标记的非特异性探针浓度(非特异性竞争实验)。于室温反应10分钟使探针与NF-κB结合,再进行PAGE。电泳后,取下凝胶制得干胶,于-70℃放射自显影12小时。②Western blot分析:5 μg细胞核蛋白提取物进行12%SDSPAGE,样品经SDS-PAGE分离后;恒压60 V转移3小时;转移后的PVD膜浸入含3%脱脂奶粉的PBST中封闭2小时;再将膜浸入1∶500稀释的鼠抗人NF-κB mAb p65中,室温反应60分钟;洗膜5次;加1∶1 500稀释的HRP-羊抗鼠IgG,室温反应60分钟。洗涤后以DAB显色试剂盒显色,终止反应,阴干后拍照。
在诸城市龙都街道大源社区,集林木种苗栽植、绿化工程施工、现代林业、旅游观光于一体的生态园林公司——大源园林生态园,现已建成7200多亩。该项目是由山东大源建设集团投资5亿元,与大黑龙沟等6个自然村联合,采用“企业+社区+合作社+农户”的模式规划建设,整体占地10000亩。目前,该园区可提供60多个品种的瓜果采摘,带动当地400多名老百姓就业,全年约接待游客20万人次。让林区变景区、田园变公园,农产品变旅游商品,大源园林生态园实现了园区旅游收益和农户采摘收益的“双丰收”,构建起“企业+农户”“林业+旅游”和谐发展的新局面。
1.3统计学分析 应用SPSS13.0软件包进行统计处理,计量资料用
2 结果
2.1MBL对DC表面标志表达的影响 流式细胞仪分析(图1)发现,与常规组相比较,MBL刺激组中CD83+、CD86+和MHC-DR+DC的数量均显著增多,加入抗MBL pAb,MBL的刺激效应消失。
2.2MBL以Ca2+依赖方式结合imDC FCM分析表明,MBL在无Ca2+、或含EDTA的结合缓冲液中几乎不与imDC细胞结合,而在分别含1 mmol/L、5 mmol/L的结合缓冲液中,MBL与imDC的结合逐渐增强,呈Ca2+浓度依赖关系(图2)。
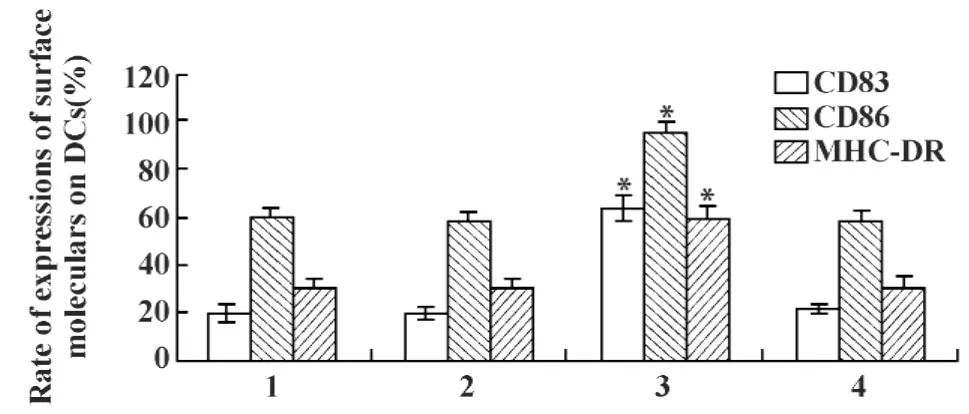
图1 不同刺激剂对DC表型的影响Fig.1 Effects of different stimulators on the phenotypes of DCs
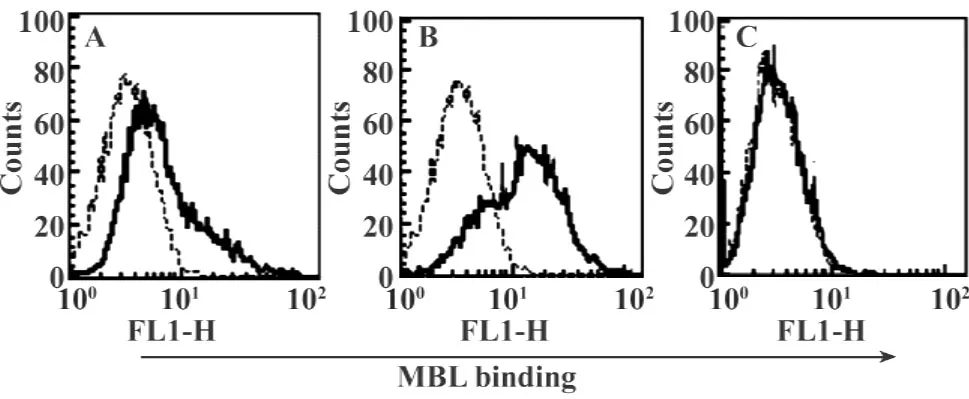
图2 MBL以Ca2+依赖方式结合imDCFig.2 Ca2+-dependent binding of MBL with imDCs
2.3MBL对imDC TLR2、TLR4 mRNA表达的影响用1%琼脂糖凝胶电泳鉴定总RNA显示28 S和18 S rRNA两条清晰带,紫外分光光度法测定总RNA样品OD260/OD280nm比值均大于1.8,表明提取的总RNA均具有较高的纯度和完整性。RT-PCR结果(图3)显示:PCR扩增产物为预期大小;随着DC的成熟,TLR2、TLR4 mRNA表达减少(Lane 2),与常规组相比,MBL组减少的更加明显(Lane 4),加入抗MBL pAb,MBL的作用效应消失(Lane 5)。
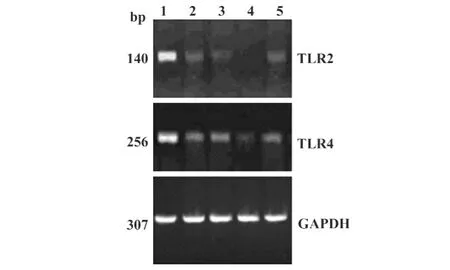
图3 MBL减弱imDC TLR2、TLR4 mRNA表达Fig.3 The decreased mRNA expressions of TLR2 and TLR4 in DCs by MBL
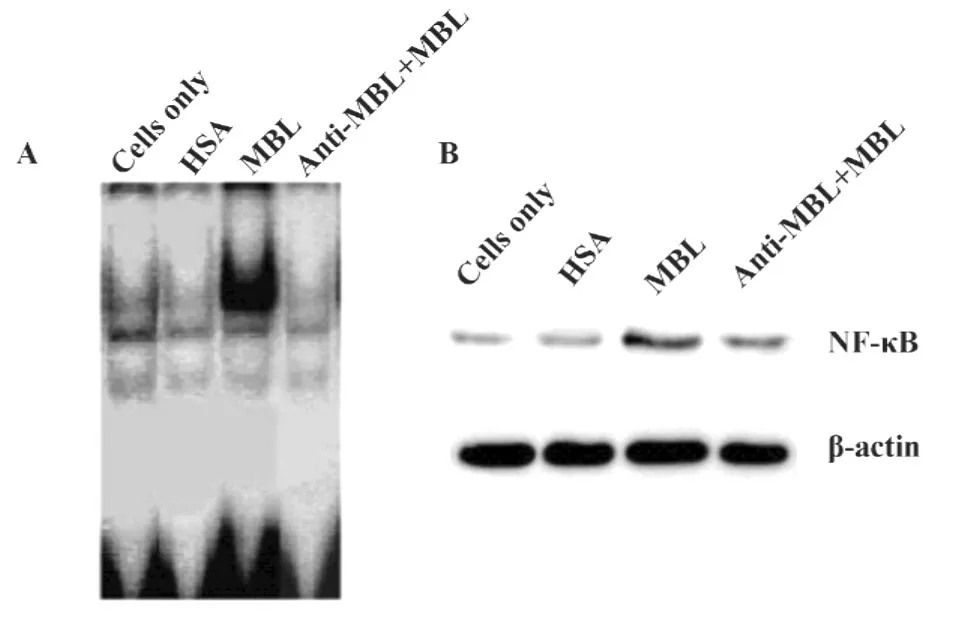
图4 MBL增强DCs NF-κB活性Fig.4 MBL enhancement of NF-κB activity in DCs
2.4MBL促进NF-κB活性 为进一步探讨MBL影响DC成熟的有关信号通路,我们又检测了MBL对核转录因子NF-κB的活性影响。EMSA结果显示:与常规组相比,MBL刺激能够显著增加imDC核转录因子NF-κB的DNA结合能力(图4A),加入抗MBL pAb,MBL的作用效应消失;Western blot结果进一步证明,MBL刺激能够显著增加imDC核转录因子NF-κB由胞浆向胞核转位,加入抗MBL pAb,MBL的作用效应消失(图4B)。
3 讨论
作为专职APC,DC在启动获得性免疫应答中扮演着极其重要的角色。DC可以通过其模式识别受体(Patten recognition receptor,PRR)识别多种病原体相关分子模式(Pathogen-associated molecule pattern,PAMP),诱导DC成熟。PRR诱导的DC成熟不仅增强其激活T淋巴细胞的能力,而且增强DC本身产生免疫调节相关的细胞因子。作为一类重要的PRR,TLR分子不仅能够介导对病原微生物及其产物的识别,而且能够参与获得性免疫应答,被视为联系天然免疫和获得性免疫的桥梁。已有研究资料表明,imDC高表达多种TLR分子,如TLR2、TLR4,但是随着DC的成熟,TLR分子的表达逐渐降低,对于完全成熟的 DC,一些 TLR分子(如 TLR2和TLR4)的表达几乎检测不到。本研究结果显示,与常规组相比,MBL组DC TLR2和TLR4表达减弱更加明显,提示MBL确实有诱导DC成熟的功能。
许多研究结果证实NF-κB与DC的功能密切相关,它不仅决定了DC激活初始T淋巴细胞的能力,而且与多种炎性细胞因子的分泌有关(如TNF-α、IL-1、IL-6 等)[14-16]。作为一类重要的 PRR,TLR 介导的信号通路可以激活多种转录因子的表达,其中核转录因子NF-κB对DC成熟和细胞因子产生至关重要,随着DC的成熟,NF-κB活性明显增强,TLRNF-κB信号途径是影响DC成熟的极为重要的信号通路。我们研究发现,与常规组相比,MBL明显增强NF-κB的DNA结合能力和由胞浆向胞核转位的能力,提示MBL通过NF-κB信号途径来调节DC成熟的。
综上所述,MBL能够与DC表面分子直接相互作用,减弱DC表面TLR2与TLR4的表达,增强DC NF-κB活性,提示MBL能够通过调控 TLR/TNF-κB信号通路来发挥其调节DC成熟功能。
1 Thompson C.Protein to proves to be a key link in innate immunity[J].Science,1995;269(5222):301-302.
2 张雅妮,王明永,雷 鸣 et al.MBL与THP1/CD14细胞结合特性的研究[J].现代免疫学,2008;28(2):121-125.
3 王明永,张雅妮,雷 鸣et al.MBL抑制Jurkat细胞增殖和分泌IL-2[J]. 现代免疫学,2010;30(6):448-452.
4 王明永,张雅妮,张丽芸et al.MBL与Raji细胞结合特性的研究[J].生物化学与生物物理进展,2008;35(3):335-340.
5 王明永,王凡平,郭晓芳 et al.MBL抑制白假丝酵母菌刺激THP1/CD14细胞产生TNF-α和IL-8[J].中华微生物学和免疫学杂志,2011;31(1):14-19.
6 Wang M,Chen Y,Zhang Y et al.Mannan-binding lectin directly interacts with Toll-like receptor 4 and suppresses lipopolysaccharide-induced inflammatory cytokine secretion from THP-1 cells[J].Cell Mol Immunol,2011;8(3):265-275.
7 Wang M,Zhang Y,Chen Y et al.Mannan-binding lectin regulates dendritic cell maturation and cytokine production induced by lipopolysaccharide[J].BMC Immunol,2011;12:1-10.
8 陈 月,陈政良,左大明et al.MBL对树突状细胞体外分化成熟的影响[J].细胞与分子免疫学杂志,2005;27(2):33-36.
9 Csomor E,Bajtay Z,Sándor N et al.Complement protein C1q induces maturation of human dendritic cells[J].Mol Immunol,2007;44(13):3389-3397.
10 王明永,张丽芸,张雅妮 et al.联合应用配体和单克隆抗体亲和层析纯化人血浆天然MBL蛋白[J].免疫学杂志,2008;24(2):119-122.
11 Bohlson S S,Fraser D A,Tenner A J.Complement proteins C1q and MBL are pattern recognition molecules that signal immediate and long-term protective immune functions[J].Mol Immunol,2007;44(1):33-43.
12 Visintin A,Mazzoni A,Spitzer J H et al.Regulation of Toll-like receptors in human monocytes and dendritic cells[J].J Immunol,2001;166(1):249-255.
13 Downing I,Macdonald S L,Turner M L et al.Detection of an autologous ligand for mannan-binding lectin on human B lymphocytes[J].Scand J Immunol,2005;62(6):507-514.
14 Baldwin A S.Series introduction:the transcription factor NF-kappaB and human disease[J].J Clin Invest,2001;107(1):3-6.
15 Thompson A G,O'Sullivan B J,Beamish H et al.T cells signaled by NF-kappa B-dendritic cells are sensitized not anergic to subsequent activation[J].J Immunol,2004;173(3):1671-1680.
16 Yoshimura S,Bondeson J,Foxwell B M et al.Effective antigen presentation by dendritic cells is NF-kappaB dependent:coordinate regulation of MHC,co-stimulatory molecules and cytokines[J].Int Immunol,2001;13(5):675-683.
猜你喜欢
现代仪器与医疗(2022年4期)2022-10-08
中国科技纵横(2021年24期)2021-03-02
中国生殖健康(2020年2期)2021-01-18
装备制造技术(2020年1期)2020-12-25
蚕桑通报(2020年1期)2020-07-10
中国土壤与肥料(2018年5期)2018-11-05
中成药(2018年6期)2018-07-11
北京航空航天大学学报(2017年2期)2017-11-24
中国医疗保险(2017年5期)2017-05-17
中国康复理论与实践(2015年10期)2015-12-24
