
酸性普鲁兰酶基因在地衣芽孢杆菌中的表达*
2011-12-18 11:23:02谢银珠沈微王正祥
食品与发酵工业 2011年2期
谢银珠,沈微,王正祥
(江南大学生物工程学院工业生物技术教育部重点实验室,江苏无锡,214122)
酸性普鲁兰酶基因在地衣芽孢杆菌中的表达*
谢银珠,沈微,王正祥
(江南大学生物工程学院工业生物技术教育部重点实验室,江苏无锡,214122)
根据Genbank公布的来源于Bacillus deramificans的普鲁兰酶基因突变体序列(AX203843)合成普鲁兰酶成熟肽基因。将该基因插入芽孢杆菌分泌型表达载体pHY-WZX,重组质粒转化地衣芽孢杆菌B60608,重组地衣芽孢杆菌实现普鲁兰酶分泌表达。对重组菌产普鲁兰酶的条件进行优化,以含2%药媒和8%甘油的培养基最适合普鲁兰酶表达。
普鲁兰酶,地衣芽孢杆菌,Bacillus deramificans
普鲁兰酶是一种能够专一性切开支链淀粉分支点中的α-l,6-糖苷键,从而剪下整个侧支,形成直链淀粉的脱支酶[1]。普鲁兰酶能将最小单位的支链分解,在淀粉制糖工业中与糖化酶混合使用能最大限度地利用淀粉。目前普鲁兰酶是淀粉制糖酶系中唯一一种我国尚不能自主生产的产品。国内外有关普鲁兰酶及其基因已有大量的文献报道,在这些信息中最具有应用价值的应该是来源于芽孢杆菌及其近缘种属的普鲁兰酶基因的信息,因为芽孢杆菌具有向细胞外大量分泌蛋白质的能力,同时芽孢杆菌的外源基因表达技术也比较成熟。为充分利用国外研究成果,本研究组根据已公布的来源于芽孢杆菌近缘种属的普鲁兰酶基因序列采用化学合成的方法合成了多条普鲁兰酶基因,并利用地衣芽孢杆菌为宿主进行异源表达,本文报道Bacillus deramificans一突变体基因的表达情况。
1 材料与方法
1.1 菌株和质粒
采用的普鲁兰酶基因为Bacillus deramificans普鲁兰酶基因突变体,其序列由NCBI数据库查寻获得,其登录号AX203843。普鲁兰酶基因由上海生物工程有限公司合成,合成后基因插入载体pUC18获得重组质粒 pUC-pulA。枯草芽孢杆菌杆菌(B.subtilis)WB600和地衣芽孢杆菌(B.licheniformis)B60608均由江南大学工业微生物技术教育部重点实验室保藏。芽孢杆菌表达载体pHY-WZX由江南大学工业微生物技术教育部重点实验室构建,该质粒以地衣芽孢杆菌淀粉酶生产菌控制淀粉酶表达的强启动子和信号肽控制外源基因的表达。
1.2 工具酶与试剂
PyrobestTMDNA聚合酶、T4DNA连接酶、各种限制性内切酶购于(大连)宝生物公司;PCR产物(小量)纯化试剂盒、质粒小量提取试剂盒购于上海华舜生物工程有限公司;引物由上海生物工程有限公司合成。Red-Pullulan多糖为爱尔兰Megazyme公司产品。标准普鲁兰酶酶液(1100 ASPU/mL)购于杰能科公司。其他试剂药品为进口或国产分析纯和生化试剂。
1.3 DNA操作技术
质粒DNA的提取、酶切、连接等参照文献[2]。DNA扩增在0.2 L PCR薄壁管中进行。PCR扩增条件为:94℃ 50 s,56℃ 90s,72℃ 3 min,循环30次。
1.4 普鲁兰酶基因表达载体的构建
根据普鲁兰酶基因序列,设计如下引物 Pyz01:5'-AATTACCGGAATTCGATGGGAACACGACAACGATC-3'和Pyz02:5'-AATTACCGGGTACCTTACTTTTTACCGTGGTCTGG-3',以pUC-pulA为模板用 PCR方法扩增出普鲁兰酶成熟肽编码基因pulA。所得片段用Kpn I和 EcoR I酶切后,插入经Kpn I和 EcoR I双酶切的载体 pHY-WZX[3],得到重组质粒pHY-WZX-pulA。
1.5 重组菌的构建
将1.4得到的重组质粒pHY-WZX-pulA转入枯草芽孢杆菌 WB600[4],筛选得到重组子 WB600(pHY-WZX-pulA)。从重组子 WB600(pHY-WZX-pulA)中提取重组质粒pHY-WZX-pulA转化地衣芽孢杆菌 B60608[5]。
1.6 普鲁兰酶酶活测定
普鲁兰酶酶活单位定义:在相应条件下,每分钟分解普鲁兰所释放的还原糖,其还原力相当于1 μmol葡萄糖所需的酶量,以1ASPU表示。普鲁兰酶酶活的测定参考Megazyme公司Red-Pullulan使用说明书并以杰能科公司市售普鲁兰酶(1100ASPU/mL)作为标准对照,验证测定方法。
1.7 发酵试验
种子培养基:(LB液体培养基):1%胰蛋白胨、0.5%酵母浸提物、1%NaCl。添加1.5%琼脂粉为LB固体培养基。添加10 μg/mL四环素分别用于重组枯草芽孢杆菌或重组地衣芽孢杆菌的筛选。
初始发酵培养基:4%碳源,1.5%棉籽粉,0.3%硫酸铵,0.03%CaCl2,11 mmol/L 磷酸盐。
优化后发酵培养基:8%甘油 ,2%药媒(棉籽蛋白粉),0.3%(NH4)2SO4,0.03%CaCl2,11 mmol/L磷酸盐。
2 结果与讨论
2.1 Bacillus deramificans普鲁兰酶基因的克隆和重组表达载体的构建
以Bacillus deramificans DNA为模板,Pyz 01和Pyz 02为引物,在PyrobestTMDNA聚合酶的作用下,PCR扩增普鲁兰酶成熟肽结构基因。扩增出的片段大小为2.8kb。将PCR产物纯化后用Kpn I和 EcoR I双酶切,与经同样酶切的pHY-WZX载体连接,直接连接产物转化枯草芽孢杆菌WB600,在含四环素的LB平板上得到3个转化子。其中1号转化子中重组质粒经 Kpn I和 EcoR I酶切电泳,产生2.8kb和6.7kb大小的片段(图1),分别与普鲁兰酶基因和空载线性pHY-WZX大小一致,符合质粒 pHY-WZX-pulA的基本特征(图2)。
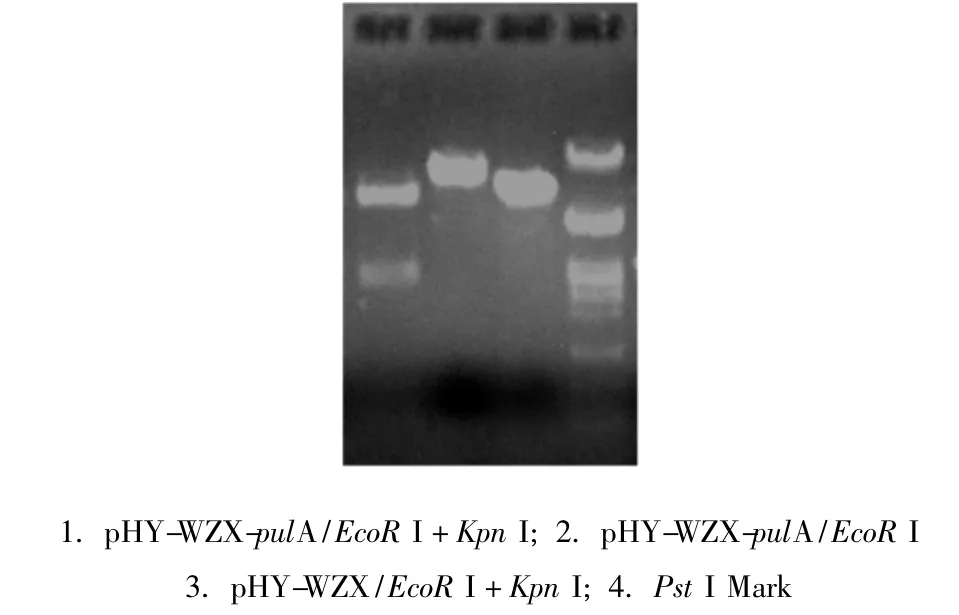
图1 重组质粒pHY-WZX-pulA酶切验证
2.2 普鲁兰酶基因的异源表达
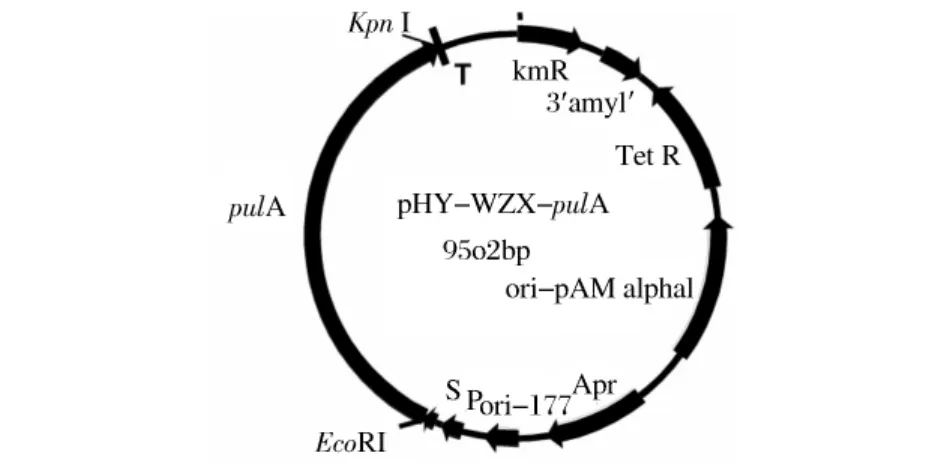
图2 重组质粒pHY-WZX-pulA
从1.5中获得的重组枯草芽孢杆菌WB600(pHY-WZX-pulA)中大量提取重组质粒pHY-WZX-pulA,用原生质体转化法转化地衣芽孢杆菌B60608。重组菌B60608接种含10 μg/mL四环素的LB液体培养基,42℃振荡培养15h后,取5 mL菌液转入发酵培养基培养一定时间,测定酶活。结果表明,含重组质粒pHY-WZX-pulA的重组地衣芽孢杆菌成功表达出有普鲁兰降解活性的普鲁兰酶,普鲁兰酶成功分泌到培养液中,培养液酶活为0.12 ASPU/mL(发酵液)而含空质粒pHY-WZX的重组地衣芽孢杆菌培养液不具有普鲁兰降解活性。
2.3 发酵优化
2.3.1 最适碳源
分别以乳糖、甘油、可溶性淀粉、糯米淀粉、马铃薯淀粉、木糖、葡萄糖为发酵培养基中的主要碳源,在同一条件下进行发酵,检测重组菌B60608产酶情况,结果见图3。
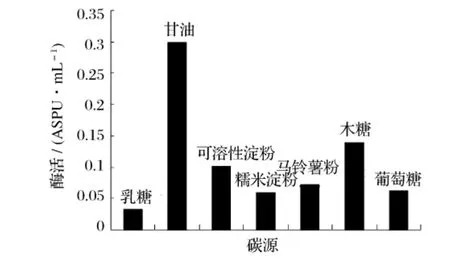
图3 不同碳源对产普鲁兰酶活力的影响
图3 表明,不同碳源对重组菌B60608产酶影响较大,甘油为产酶最佳碳源。以甘油为碳源,采用不同的加量,保持其他条件不变进行摇瓶发酵实验,结果见图4。由图4可知,培养基中最适的甘油含量为8%。
2.3.2 最适有机氮源
选取几种不同的有机氮源作为培养基中的唯一有机氮源,在同一条件下进行发酵,测定不同有机氮源对重组菌B60608发酵产酶的影响,产酶结果见图5。由图5可知,以药媒作为氮源时所获酶活最高。以药媒为唯一氮源,考察不同药媒加量对产酶影响,结果见图6。从图6中可以得出,2%药媒为发酵培养基中最适有机氮源。

图4 甘油添加量对产普鲁兰酶活力的影响
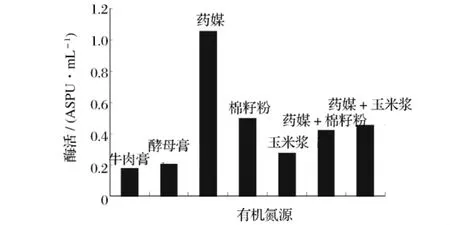
图5 不同有机氮源对产普鲁兰酶活力的影响
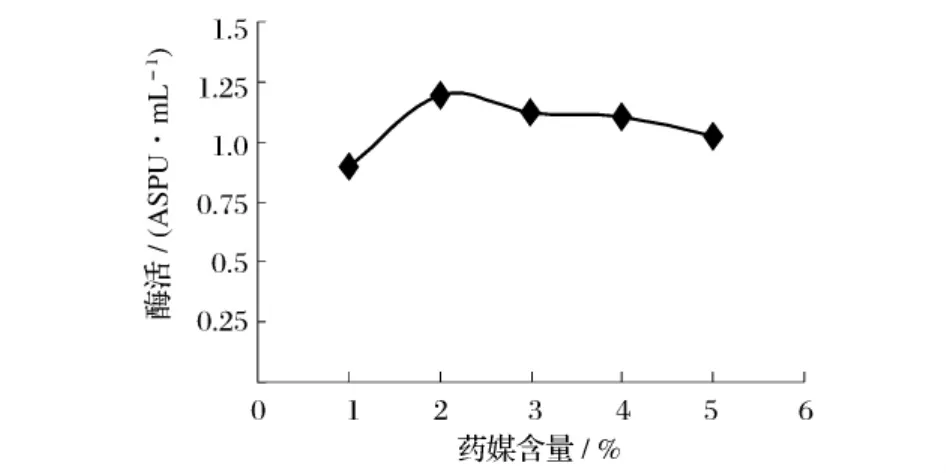
图6 药煤含量对产普鲁兰酶活力的影响
2.3.3 最适无机氮源
选取不同的无机氮源,在同一条件下进行发酵,考察其对产酶的影响,结果见图7。由图7可知,(NH4)2SO4对产酶具有促进效果,选取(NH4)2SO4为无机氮源。以(NH4)2SO4作为无机氮源,考察不同加量的(NH4)2SO4对产酶影响,结果见图8。由图8可知,0.3%(NH4)2SO4为发酵培养基最适无机氮加量。
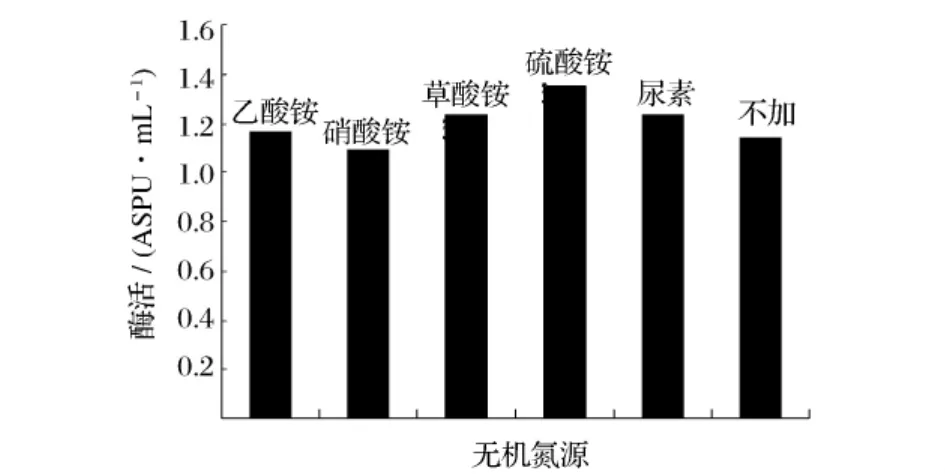
图7 不同无机氮源对产普鲁兰酶活力的影响
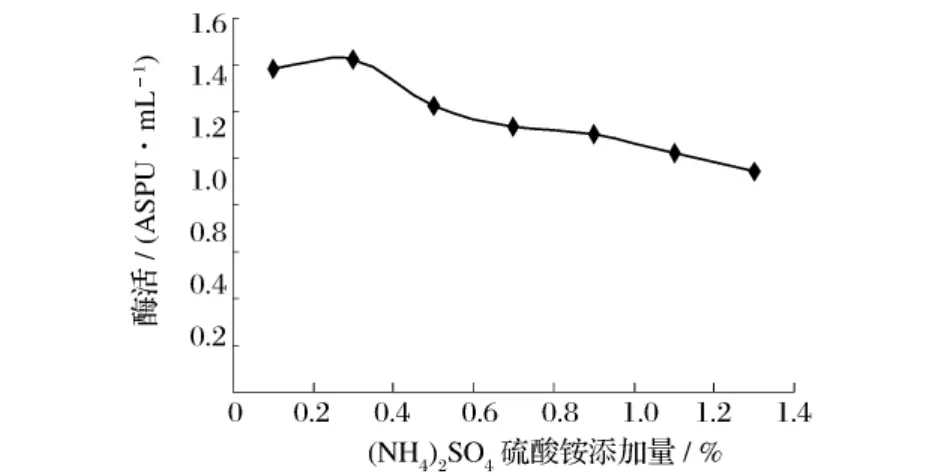
图8 (NH4)2SO4浓度对产普鲁兰酶活力的影响
3 讨论
本研究组根据国外专利合成了多条来源于芽孢杆菌及其近缘种属的普鲁兰酶基因,并以大肠杆菌和地衣芽孢杆菌为宿主进行异源表达,结果发现这些基因中只有少数能表达出明显的普鲁兰降解活性(另文发表),其中诺维信公司和杰能科公司公布的来源于Bacillus deramificans和Bacillus acidopullulyticus的普鲁兰酶基因的表达产物具有较高的普鲁兰降解活性。本文报道的是研究来源于Bacillus deramificans的普鲁兰酶基因的一个突变体在地衣芽孢杆菌中的表达情况,该基因编码产物为928个氨基酸残基的成熟肽蛋白,分子量约10万,经条件优化后摇瓶发酵水平为1.5 ASPU/mL,酶活存在于培养液中。据了解美国杰能科公司等也是采用重组地衣芽孢杆菌生产普鲁兰酶,而普鲁兰酶基因也是Bacillus deramificans来源的突变体,其发酵产酶水平估计在1000 ASPU/mL以上。蛋白电泳显示,含重组质粒 pHY-WZX-pulA的重组地衣芽孢杆菌与含pHY-WZX空质粒的地衣芽孢杆菌的蛋白谱在10万左右未见明显差异,这说明重组菌普鲁兰酶蛋白表达量还很低,提高蛋白表达量是提高普鲁兰酶表达水平的关键技术。
4 结论
本研究利用芽孢杆菌分泌型表达载体pHY-WZX研究了来源于Bacillus deramificans的普鲁兰酶的一个突变体基因pulA的表达方法。在地衣芽孢杆菌淀粉酶基因的启动子和信号肽控制下pulA成功实现分泌表达,表达产物具有Red-Pullulan多糖降解活性并且全部分泌到培养液中。对重组菌发酵条件进行优化,获得最优化的培养基组成;碳源为8%甘油,氮源为2%药媒及0.3%的硫酸铵。
[1] Nair SU,Singhal RS,Kamat MY.Induction of pullulanase production in Bacillus cereus FDTA-13[J].Bioresource Technology,2007,98(4):856-859.
[2] 诸葛健,王正祥.工业微生物实验技术手册[M].北京:中国轻工业出版社,1994.
[3] Niu D,Wang ZX.Development of a pair of bifunctional expression vectors for Escherichia coli and Bacillus licheniformis[J].Journal of Industrial Microbiology and Biotechnology,2007,34(5):357-362.
[4] Imanaka T,Tanaka T,Tsunekawa H,et al.Cloning of the genes for penicillinase,penP and penI,of Bacillus licheniformis in some vector plasmids and their expression in Escherichia coli,Bacillus subtilis,and Bacillus licheniformis[J].Journal of Bacteriology,1981,147(3):776 -786.
[5] 莫静燕,陈献忠,王正祥.地衣芽孢杆菌原生质体的制备、再生及转化研究[J].生物技术,2009,19(5):75-77.
Expression of the Acid Pullulanase in Bacillus licheniformis
Xie Yin-zhu,Shen Wei,Wang Zheng-xiang
(Key Laboratory of Industrial Biotechnology of Ministry of Education and School of Biotechnology,Jiangnan University,Wuxi 214122,China)
According to the sequence of pullulanase gene from Bacillus deramificans(NCBI accession number:AX203843),the gene encoding mature peptide of pulluanase was synthesized and designated pulA.The pulA was amplified by the method of PCR and cloned into the expression vector pHY-WZX,yielding hybrid plasmid pHYWZX-pulA.Subsequently,pHY-WZX-pulA was introduced into Bacillus licheniformis B60608.Active pullulanase was expressed by recombiant B.licheniformis and secreted into medium.Culture condition of recombinant B.licheniformis were optimized for production of pullulanase.The optimized medium consists of 2%cotton seed protein and 8%of glycerol.
pullulanase,Bacillus licheniformis,Bacillus deramificans
硕士研究生(王正祥教授为通讯作者,E-mail:zxwang@jiangnan.edu.cn)。
*国家863高技术研究发展计划(2006AA10Z307)
2010-08-01,改回日期:2010-11-30
猜你喜欢
食品与药品(2020年1期)2020-03-10 03:25:00
山东工业技术(2016年15期)2016-12-01 05:31:50
中国酿造(2016年12期)2016-03-01 03:08:19
中国酿造(2016年12期)2016-03-01 03:08:11
应用海洋学学报(2015年3期)2015-11-22 07:39:30
中国酿造(2014年9期)2014-03-11 20:21:03
食品工业科技(2014年13期)2014-03-11 18:16:48
食品工业科技(2014年9期)2014-03-11 18:15:28
中国粮油学报(2014年8期)2014-02-06 01:34:10
中国食用菌(2013年2期)2013-11-21 03:37:52
