
普鲁兰糖生物合成和分子量调控机制的研究进展
2020-03-10 03:25:00刁梦奇张金华张林军王凤山凌沛学
食品与药品 2020年1期
刘 飞,刁梦奇,张金华,张林军,袁 超,王凤山,凌沛学
(1.山东大学药学院,山东 济南 250012;2.山东省药学科学院 山东省生物药物重点实验室 山东省多糖类药物工程实验室 多糖类药物发酵与精制国家地方联合工程实验室 博士后科研工作站,山东 济南 250101;3.山东福瑞达医药集团有限公司,山东 济南 250101)
普鲁兰糖是由出芽短梗霉发酵产生的一种胞外多糖,又称普鲁兰多糖、短梗霉多糖、茁霉多糖等。Bernier最先将普鲁兰糖分离并解析其结构[1],后将其命名为“普鲁兰糖”[2]。当前对普鲁兰糖的研究大多是围绕其应用与筛选,而有关普鲁兰糖分子合成途径及分子量调控方面的研究较少。
1 普鲁兰糖的结构与性质
研究发现[3-4]普鲁兰糖是α-1,4糖苷键连接的麦芽三糖重复单位通过α-1,6糖苷键聚合而成的直链状多糖(图1),分子量范围2×104~2×106,分子量受到菌种及培养条件等多重因素影响[5-6]。普鲁兰糖是一种非离子型、非吸湿性的聚合物,具有良好的可塑性、稳定性、成膜性和安全性,其黏度相对低于其他聚合物[7]。由于这些独特的性质,普鲁兰糖在食品、药物和生物医学等领域均有广泛的应用[8-11]。
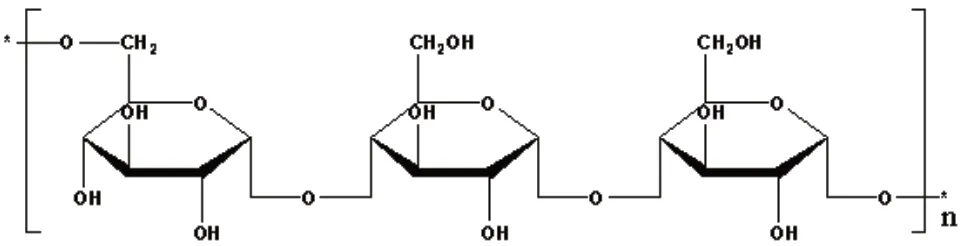
图1 普鲁兰糖的基本结构
2 普鲁兰糖分子合成途径的研究进展
2.1 普鲁兰糖的分子合成关键步骤
目前,有关普鲁兰糖合成的具体分子机制尚未完全清楚。在普鲁兰糖的生物合成过程中约有5种酶参与(表1),尿苷二磷酸葡萄糖焦磷酸化酶(UDPG-焦磷酸化酶)、α-磷酸葡萄糖变位酶和葡萄糖基转移酶被认为是其中的关键酶[12]。从碳源开始至最后产生普鲁兰糖,可将其分为三大关键步骤。
2.1.1 从碳源初始底物到UDPG 出芽短梗霉利用碳源合成普鲁兰糖,Duan等[13]研究发现,虽然普鲁兰糖合成可利用多种碳源,如甘露糖、蔗糖、麦芽糖、果糖、半乳糖、木糖等,但在同等条件下,当碳源为葡萄糖时,普鲁兰糖的产量最大。在转化过程中,己糖激酶等负责将碳源转化为6-磷酸-葡萄糖,α-磷酸葡萄糖变位酶催化6-磷酸-葡萄糖形成1-磷酸-葡萄糖[13]。普鲁兰糖前体UDP-葡萄糖是由UDPG-焦磷酸化酶将1-磷酸葡萄糖和UDP催化为UDP-葡萄糖和焦磷酸(pyrophosphoric acid,PPi),Shingel等[14]研究表明,UDPG不能由ADPG(adenosine diphosphate glucose)取代,表明普鲁兰糖起源于UDPG,在此反应中,ATP(adenosinetriphosphate)和NADPH(nicotinamide adenine dinucleotide phosphate)作为辅助因子是必需的[13-14]。UDP-葡萄糖是普鲁兰糖合成的重要前体[14]。

表1 普鲁兰糖合成过程中相关酶
2.1.2 从UDPG到异麦芽糖 Shingel等[14]研究表明,含葡萄糖的脂质中间体在普鲁兰糖的形成中起着至关重要的作用:首先UDP-葡萄糖介导的D-葡萄糖残基通过磷酸酯桥连接到脂质分子(lipid molecule,LPh)上,进一步添加UDP-葡萄糖的D-葡萄糖残基,得到脂质连接的异麦芽糖;接下来,异麦芽糖基与脂质连接的葡萄糖参与反应,生成异戊酰基残基,异戊酰基残基可聚合到普鲁兰糖链中。Hayashi等[15]研究表明葡萄糖基转移酶在从麦芽糖(Glc-α-1→4-Glc)产生潘糖(Glc-α-1→6-Glc-α-1→4Glc)及异麦芽糖(Glc-α-1→6-Glc)中发挥重要功能。同时Duan等[13]研究表明,葡萄糖基转移酶活性水平越高,普鲁兰糖的产量越高,但其发挥功能具体机制尚不明确。
2.1.3 从异麦芽糖到普鲁兰糖链 有文献报道,从异麦芽糖形成普鲁兰糖链的过程是在葡萄糖基转移酶的作用下完成的[12,16]。但是,Kang等[17]构建了破坏普鲁兰合酶基因的突变菌株,发现其可产生纯的β-葡聚糖胞外多糖,而不能产生普鲁兰糖。Chi等[18]也发现破坏菌株中的普鲁兰合成酶可降低普鲁兰糖产量,暗示在从异麦芽糖形成过程中也可能是普鲁兰合酶在发挥作用[4]。
已知己糖激酶、UDPG-焦磷酸化酶、α-磷酸葡萄糖变位酶、葡萄糖基转移酶、普鲁兰合酶在普鲁兰糖的形成过程中有重要功能,本文研究人员根据文献绘制了普鲁兰糖的合成途径(见图2):己糖激酶催化葡萄糖形成葡萄糖-6-磷酸;葡萄糖-6-磷酸通过-磷酸葡萄糖变位酶形成葡萄糖-1-磷酸;葡萄糖-1-磷酸在UDPG-焦磷酸化酶作用下生产二磷酸尿苷葡糖;然后通过葡萄糖基转移酶与脂质体生产连接脂质的葡萄糖;连接脂质的葡萄糖通过葡萄糖基转移酶和普鲁兰合酶最终形成普鲁兰糖。但其具体作用机制尚未得到明确的阐释,这也为下一步的研究提供了目标。只有明确其作用机制,才能有针对性地控制菌种和产量等,更好地应用于工业生产和科研工作。
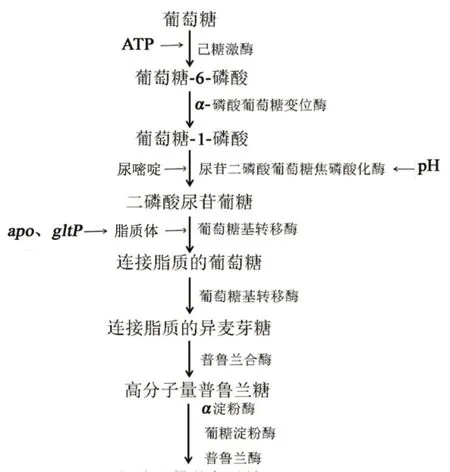
图2 普鲁兰糖合成途径
2.1.4 其他酶及化合物对普鲁兰糖合成的影响 除上述参与合成的酶外,其他酶及物质也会影响普鲁兰糖的合成。郭建等[19]发现,整合过表达载脂蛋白基因apo或gltP能增加多糖产量,而敲除apo或gltP则使多糖产量降低。作者推测可能因为apo和gltP调控脂类物质合成,影响普鲁兰糖合成所需基本单元糖脂中间体lipid-葡萄糖生成,或是导致某些脂类物质作为反式作用因子强化了UDPG合成,导致磷酸葡萄糖变位酶、UDPG焦磷酸化酶和葡萄糖基转移酶的酶的活性增加,最终影响普鲁兰糖合成。研究[20-21]出芽短梗霉菌株中过表达透明颤菌血红蛋白(Vitreoscillahemoglobin,VHb)和黄素血红蛋白(flavohemoglobin,FHB),发现菌株携氧能力明显增强,负责葡萄糖运输、糖酵解、NADH代谢、ATP产生等基因的转录水平提高,显著提高了普鲁兰糖产量。可能因为普鲁兰糖的合成是一个消耗ATP的过程,与此同时,在大肠杆菌中过表达VHb可提高ATP的生产效率。由此推测,改变在普鲁兰糖合成过程中调控中间体合成的酶活性,也可间接调控普鲁兰糖的产量。如,改变其他调控脂质、ATP合成等酶的活性,也可能会对普鲁兰糖的产量产生影响,为改造菌种提供了一个新思路。
此外,改变菌种的培养环境也可调节普鲁兰糖的产量。Sheng等[22]研究发现,在培养时添加腺嘌呤、鸟嘌呤和胞嘧啶虽然增加了生物量,但是并未促进普鲁兰糖产生,而增加尿嘧啶可增加普鲁兰糖的积累量。尿嘧啶的添加时间很重要:在发酵时间0 h时添加,尿嘧啶仅作为为氮源刺激细胞生长,而普鲁兰糖的产量减少;但是在48 h添加,普鲁兰糖产量增加。本文研究人员推测可能因为尿嘧啶是UDP-葡萄糖的前体,UDP-葡萄糖是普鲁兰糖合成的关键因素,添加的尿嘧啶可能提供了额外的葡萄糖基供体。
Pan等[23]通过实验发现,在pH为5.5时可得到最高普鲁兰糖产量和最高UDP-葡萄糖焦磷酸化酶的活性,pH过高或过低会影响普鲁兰糖产量和UDP-葡萄糖焦磷酸化酶活性。Wang等[24]研究发现,在培养基中添加CuSO4,能增加关键生物合成酶的活性,使细胞内ATP维持在较高水平,普鲁兰糖的分泌速率加快,所有这些均有助于提高普鲁兰糖产量。由此可见,在不清楚如何调控普鲁兰糖的合成途径时,也可通过改变其培养环境从而提高产量。
2.1.5 副产物对普鲁兰糖合成的影响 在普鲁兰糖的生产过程中,也会产生重油、黑色素等副产物。重油是普鲁兰糖所产脂类物质的主要成分。有文献研究表明,普鲁兰糖产量和重油产量呈正相关,研究者推测可能因为重油参与普鲁兰糖合成所需基本单元糖脂中间体 lipid-葡萄糖的生成[19]。但是Price等[25]发现过度强化的重油合成能力能够抑制普鲁兰糖的生成,由此看来并非重油合成能力越强普鲁兰糖合成能力越强。而在普鲁兰糖的生产过程中产生的黑色素,会给最后的产物带来较深的颜色,导致额外的脱色过程,增加成本[26]。Zheng等[27]认为普鲁兰糖与黑色素之间的关系是正相关的,但是Guo等[28]却得出了相反的结果。因此,黑色素与普鲁兰糖之间的关系需研究者进行更多、更深入的探究。探索副产物是否影响普鲁兰糖的产量,以及影响的具体机制,有助于进一步了解普鲁兰糖的合成和调控。
2.2 普鲁兰糖分子量的调控机制和影响因素
普鲁兰糖的实际应用除了受到合成产量的影响之外,还受到其分子量的制约,而其分子量也受到多种条件的影响。
普鲁兰酶是一种脱支酶,可特异性水解普鲁兰糖中的α-1,6糖苷键[29]。根据文献报道基于底物特异性和终产物,可将其分为5类[30]:(i)I型普鲁兰酶特异性水解普鲁兰糖中的α-1,6糖苷键以产生麦芽三糖[31-32];(ii)II型普鲁兰酶(amylopullulanase)攻击普鲁兰糖中的α-1,6糖苷键,产生葡萄糖、麦芽糖和麦芽三糖的混合物;(iii)I型普鲁兰水解酶(neopullulanase)攻击普鲁兰糖中的α-1,4键,产生潘糖[33];(iv)II型普鲁兰水解酶(isopullulanase)攻击普鲁兰糖中的α-1,4连接以产生异葡糖基麦芽糖,产生异潘糖[34];(v)III型普鲁兰水解酶攻击普鲁兰糖中的α-1,6以及α-1,4键,形成麦芽三糖、潘糖、麦芽糖和葡萄糖的混合物。
Liu等[35]研究确认α-淀粉酶、葡糖淀粉酶和普鲁兰酶在高分子量普鲁兰糖的生物合成中起着重要作用。删除它们的单个基因后,产生的普鲁兰糖的分子量都能大大增加,但产量下降。基因回补后,普鲁兰糖的产量恢复,分子量回降。该实验结果表明,α-淀粉酶,葡糖淀粉酶和普鲁兰酶可调节普鲁兰糖的分子量。普鲁兰糖分子量的提高会使发酵液黏度变大,影响溶氧和传质,导致产量下降。
据报道,在普鲁兰糖合成的过程中,过高的碳源会抑制普鲁兰糖合成。Wang等[36]删除菌种中的CREA基因(编码葡萄糖抑制因子)后,普鲁兰糖产量增加,分子量降低;进行CREA基因回补后,普鲁兰糖产量减少。同时,在删除CREA基因菌种中检测到编码-淀粉酶,葡糖淀粉酶和普鲁兰酶的基因转录水平显著提高。
普鲁兰糖分子量的大小除了受到多种酶的调节外,也受其他培养条件的影响。Sheng等[37]研究发现,在培养基中添加Tween 80可显著增加普鲁兰糖产量,且0.5 %Tween 80是最佳浓度;但添加后,获得的普鲁兰糖的分子量与对照组相比明显降低,原因可能是加入Tween 80改变了细胞膜的通透性,加快了细胞膜内外的物质交换。Wang等[38]在培养基中添加3 g/L的NaCl后,使普鲁兰糖浓度提高了26.7 %,但分子量降低到原来的46.8 %。进一步研究发现,NaCl增加了α-磷酸葡萄糖变位酶、葡萄糖基转移酶、α-淀粉酶的活性及转录水平,同时增强了ATP的供应,并将细胞内UDP-葡萄糖维持在较高水平,原因可能是NaCl增加普鲁兰糖产量但降低了其分子量。
由此可见,在获得较高普鲁兰糖产量时,其分子量一般较低;当获得较高分子量的普鲁兰糖时,其产量较低。如何在提高普鲁兰糖产量的同时调控其分子量的变化,制约着其工业化大规模生产,也为进一步研究普鲁兰糖产业化研究指明了方向。
3 展望
由于普鲁兰糖独特的结构与性质,在自然界可被微生物降解利用,不会引起环境污染,在食品、化工、医药等领域具有越来越广泛的应用,进一步推动其工业化生产工艺的不断改进和提高;而如何获得高产菌种、减少副产物生成、降低生产成本则成为研究的关键。因此,阐明普鲁兰糖合成途径的各个步骤及各种酶在其中的作用机制、优化其培养条件、探索更多可进行调控的酶及物质,是工业化高效生产的关键与基础,也有助于普鲁兰糖产品更快的推向市场。
猜你喜欢
保健与生活(2022年13期)2022-07-06 21:16:02
保健与生活(2022年11期)2022-06-09 22:24:39
生物学通报(2020年10期)2020-08-13 08:52:24
阅读(低年级)(2019年11期)2019-12-27 04:16:34
现代食品科技(2018年10期)2018-11-06 12:54:44
作文与考试·小学高年级版(2017年24期)2017-12-14 22:00:01
保健与生活(2017年5期)2017-04-14 07:45:59
国际妇产科学杂志(2016年2期)2016-06-16 02:10:22
食品工业科技(2014年13期)2014-03-11 18:16:48
中国粮油学报(2014年8期)2014-02-06 01:34:10
