CO2阶跃变化对茄子动态光合特征参数的影响
2011-10-11 09:21:36薛占军高志奎吕桂云高荣孚
Biophysics Reports 2011年2期
薛占军, 高志奎, 王 梅, 吕桂云, 杜 爽, 高荣孚
1.河北农业大学园艺学院,保定 071001;
2.北京林业大学生物科学与技术学院,北京 100083
CO2阶跃变化对茄子动态光合特征参数的影响
薛占军1, 高志奎1, 王 梅1, 吕桂云1, 杜 爽1, 高荣孚2
1.河北农业大学园艺学院,保定 071001;
2.北京林业大学生物科学与技术学院,北京 100083
针对CO2阶跃变化下茄子光合的振荡动态现象,采用CO2阶跃起点 (Ca1)和阶点 (Ca2)处理,以及不同光照水平 (PAR)下的CO2阶跃处理,结合光合系统反馈控制动态生化模型中传递函数的振荡单元,分析了茄子动态光合的振荡特征参数。通过稳态光合参数,将茄子光合速率 (Pn)转化为羧化速率 (Vc)的动态进程,发现在较低的CO2阶跃起点 (Ca1≤180 μmol/L)下,光合系统的时间常数T较大,光合系统的阻尼系数ζ、无阻尼自然振荡频率ωn和阻尼振荡频率ωd则较小,因而Pn和Vc振荡剧烈;较高的CO2阶跃阶点 (Ca2≥900 μmol/L)下,ζ急剧减小,Pn和Vc振荡愈加剧烈;光强PAR增至900 μmol·quata/m2·s以上时,ζ急剧减小,Pn和Vc振荡也愈加剧烈。综合分析可见,发生光合振荡的条件是较低的ζ和较大的T;相反,不发生光合振荡的条件是较高的ζ和较小的T;但是,发生光合振荡的临界条件需要在中高的ζ下配合较大 (甚至更大)的T。本试验中,能够引起茄子光合振荡的CO2阶跃起点、阶点和光强临界量分别选定为 300 μmol/L、 900 μmol/L、 500 μmol·quata/m2·s。 据此计算的 CO2阶跃扰动强度 Dc与茄子光合振荡系统的阻尼系数ζ之间呈指数极显著负相关关系 (r=-0.858**)。
动态光合;振颤;CO2阶跃变化;阻尼系数;振荡频率;时间常数
引 言
20世纪50年代就已有关于叶片光合动态变化的研究报道[1,2]指出,从低CO2或低光强转到饱和CO2或饱和光强时,植物叶片的动态光合呈现振荡现象[3~6]。温室茄子叶片光合非稳态性也受到田间环境因子的影响[7]。虽然关于动态光合的振荡机制仍无定论,但研究的焦点更多地倾向于光合系统的反馈控制。
Laisk等从反馈机制角度出发,结合rubisco酶动力学响应,先后尝试了以碳转运、电子传递链或无机磷 (Pi)再生供应建立的动态光合生化模型[8~12]。高志奎等针对CO2阶跃变化下茄子光合振荡动态,依据经典的光合系统酶触反应动力学[13],采用经典的控制系统传递函数分析手段[14],构建了光合系统反馈控制动态生化模型[15]。无论如何,振荡式的动态光合过程必然涉及光合系统中叶绿体的快速响应机制,其中调节与PSI环式电子流密切相关的ATP/NADPH输出速率和改变捕光天线色素到电子传递的能流效率尤为重要[16,17]。同时,长期高浓度CO2处理下植物叶片光合能力明显下调,其下调的内在限制机制研究,已由CO2的羧化固定转向了与光系统中捕光天线色素和电子传递相关的组份上[18];而短期高浓度CO2处理下,植物叶片光合速率的改变则主要受核酮糖-1,5-二磷酸 (ribulose-1,5-bisphosphate,RuBP)浓度或核酮糖-1,5-二磷酸羧化/加氧酶 (ribulose-1,5-bisphosphate carboxylase/oxygenase,Rubisco)的数量与活性的影响[19]。
因此,本研究以日光温室茄子 (Solanum melongena L.)为试材,在不同光照水平下进行CO2浓度阶跃变化处理,结合光合系统反馈控制动态生化模型中传递函数的振荡单元,分析了茄子动态光合的振荡特征参数,以期明确光合动态受CO2阶跃扰动的程度及其影响因子,并充实该动态生化模型的研究,为动态光合的深入研究奠定基础。
材料与方法
试验材料
植物材料为茄子。于2006年12月1日和2010年7月15日在河北农业大学日光温室播种育苗,2007年3月1日和2010年8月15日栽植,室内冠层顶部平均光强为室外的78.2%,通过放风调控,保持晴天白天室内温度在28~30℃且≯32℃,夜间温度为12~10℃且≮10℃,其它农艺管理按照常规进行。分别于2007年4~5月和2010年9~10月进行研究测定。动态光合测定
采用Ciras-2便携式光合系统 (英国PP Systems公司)进行动态光合及其相关参数测定。为了增强Ciras-2的动态响应测试性能,将光合叶室与CIRAS-2主机的连接气管从原有的130 cm改为35 cm,以时间间隔为2.27 s自动记录数据300 s。记录数据包括净光合速率(Pn)、气孔导度 (Gs)、蒸腾速率 (E)、光合有效辐射的光量子通量密度 (PAR)、气温(Ta)、叶温 (Tl)、环境二氧化碳浓度 (Ca)和叶肉细胞间隙二氧化碳浓度 (Ci)等参数。采用植株顶部第3~4片叶进行连体3次重复测定。
光量子通量密度 (PAR)和叶温 (Tl)的调节采用Ciras-2内置系统进行。CO2浓度的调节采用液体CO2与碱石灰相结合配气气袋方式获得。
CO2浓度阶跃变化处理
处理Ⅰ:在 PAR=(1500±9)μmol·quata/m2·s和 Tl=(29.9±0.2)℃下,CO2浓度 (Ca)阶跃变化为:从阶跃起点Ca1突然转换为阶跃阶点Ca2。阶跃起点的处理方法是,当进气通道Ca1分别送入5、180、360、540、720和900 μmol/L CO2的气体600 s(待Pn稳定)后,突然转换为通道Ca2气体,其Ca2为1100 μmol/L。
处理Ⅱ:在 PAR=(1500±10)μmol·quata/m2·s和 Tl=(29.8±0.6)℃下,Ca阶跃阶点的处理方法是,当进气通道Ca1送入8 μmol/L CO2的气体600 s(待Pn稳定)后,突然转换为通道Ca2气体,其 Ca2分别为 300、600、900、1200、1500 和 1800 μmol/L。
处理Ⅲ:在 PAR 分别为 300、600、900、1200、1500、1800 μmol·quata/m2·s 和 Tl=(29.9±0.1)℃下,Ca阶跃变化处理方法是当进气通道Ca1送入15 μmol/L CO2的气体600 s(待Pn稳定)后,突然转换为通道Ca2气体,其Ca2为1200 μmol/L。
稳态光合参数计算
羧化速率Vc参考von Caemmerer[13]的方法,计算式为Vc=(Pn+Rd)/(1-Γ*/Ci)。其中,Pn为实测净光合速率,Rd为光下暗呼吸速率,Γ*为叶绿体CO2分压下的CO2补偿点,Ci为胞间CO2浓度。这里,参考von Caemmerer方法[13]中的式2.33和表2.3计算了下列参数:Γ*=38.6×1.37(Tl-25)/10,Rd=0.02×80×2.46(Tl-25)/10。另外,还计算了 rubisco 酶的羧化动力学米氏常数 Kc=260×2.24(Tl-25)/10和加氧动力学米氏常数 Ko=179×1.63(Tl-25)/10。
动态光合振荡特征参数的计算
光合系统反馈控制动态生化模型结构以卡尔文环的rubisco羧化/加氧接触反应的酶动力学进程作为核心、以PGA还原和接续的RuBP再生的多级过程高度简化为复合酶接触反应的酶动力学进程为反馈而构成[15]。其表达式为G:
G=k0(s+ωb1)(s+ωb2)(s2+2ζbωb3s+ωb32)/〔(s+ωa1)(s+ωa2)(s+ωa3)(s+ωa4)(s+ωa5)(s+ωa6)(s+ωa7)(s+ωa8)(s+ωa9)
(s2+2ζaωa10s+ωa102)〕
这里,s为经Laplace变换的复变量,ω为无阻尼自然频率、ζ为系统的阻尼系数。其中,ωb1=k8,ωb2=0.8333k9,ωb3=(k4k6)0.5,2ζb=(k4Ko+k4)/〔2(k4k6)0.5〕;ωa1、ωa2、ωa3、ωa4、ωa5、ωa6、ωa7、ωa8、ωa9、ωa10和 ζa分别为 k1、k2、……、k11与 Kc、Ko、Kr、O 组成的系数群。其中,Kc=(k3+k2)/k1,Ko=(k6+k5)/k4,Kr=(k11+k10)/k9。
从CO2阶跃响应传递函数中的振荡单元ωn2/(s2+2ζωns+ωn2),可以计算茄子动态光合的振荡特征参数。其中,系统的阻尼系数 ζ={(lnσ)2/〔π2+(lnσ)2〕}0.5、无阻尼自然振荡频率 ωn=4/(ζts)、阻尼振荡频率ωd=ωn(1-ζ2)0.5和时间常数T=1/ωn。超调量σ为振荡进程中光合速率最大峰值超过稳态值的比例;调整时间ts为振荡进程中光合速率开始达到并保持在允许误差(2%或5%)范围之内 (即稳态值)所需要的时间。
为了探讨CO2阶跃变化对茄子光合振荡程度的影响,引入CO2阶跃扰动强度的概念。该概念表达为,Dc=(Ca2-Ca1)/Ca2·(Ca1o/Ca1)·(Ca2/Ca2o)·(PAR/PARo)。其中,Ca1为 CO2阶跃的起点,Ca2为CO2阶跃的阶点,PAR为光合动态进程中的光强,Ca1o、Ca2o、PARo分别为Ca1、Ca2、PAR能够引起光合振荡的临界量。
结果与分析
CO2阶跃起点对茄子光合动态响应特征的影响
通过稳态光合参数,将处理Ⅰ中CO2阶跃变化下茄子光合速率 (Pn)转化为羧化速率(Vc)的动态进程,可见二者明显受CO2阶跃的起点影响 (图1)。从图1可见,较低的CO2阶跃起点 (Ca1=5或180 μmol/L),引起Pn和Vc振荡现象较为强烈;而随着CO2阶跃起点的升高 (Ca1=540、720或900 μmol/L),Pn和Vc振荡现象趋于不明显。

图1 CO2阶跃起点Ca1对茄子光合速率Pn(A)和羧化速率Vc(B)动态进程的影响Fig.1 Effect of CO2step starting pointCa1on the dynamic course of net photosynthesis ratePn(A)and carboxylation rateVc(B)in eggplant
将图1中Vc的动态进程进行振荡特征分析表明,当CO2阶跃的起点较低 (Ca1=5或180 μmol/L)时,时间常数T较大,系统的阻尼系数ζ、无阻尼自然振荡频率ωn和阻尼振荡频率ωd则较小,因而振荡剧烈 (图2)。同时从图2可见,随着CO2阶跃起点的升高,T呈现降低趋势,且在CO2阶跃的起点Ca1超过360 μmol/L以上时趋于最小,而随着CO2阶跃起点的升高,ζ、ωn和ωd则呈现增加趋势,且在CO2阶跃的起点Ca1超过360 μmol/L以上时,ζ趋于最大,可是ωn比ωd增加得更快。

图2 CO2阶跃起点Ca1对茄子光合系统的阻尼系数ζ、无阻尼自然振荡频率ωn和阻尼振荡频率ωd(A),以及时间常数T(B)的影响Fig.2 Effect of CO2step starting pointCa1on the damp coefficient ζ,natural oscillation frequency for undamped presentωn,damped oscillation frequencyωd(A),and time constantT(B)of photosynthetic system in eggplant
CO2阶跃阶点对茄子光合动态响应特征的影响
在处理Ⅱ的CO2阶跃变化下,茄子Pn和Vc的动态进程也明显受CO2阶跃阶点的影响(图3)。从图3可见,CO2阶跃阶点较高 (Ca2>900 μmol/L),引起Pn和Vc振荡现象较为强烈;而随着CO2阶跃阶点的降低,在Ca2=900 μmol/L下,Pn和Vc振荡减缓;当Ca2≤600 μmol/L时,Pn和Vc振荡现象趋于不明显。

图3 CO2阶跃阶点Ca2对茄子光合速率Pn(A)和羧化速率Vc(B)动态进程的影响Fig.3 Effect of CO2step order pointCa2on the dynamic course of net photosynthesis ratePn(A)and carboxylation rateVc(B)in eggplant
将图3中Vc的动态进程进行振荡特征分析表明,CO2阶跃的阶点较低 (Ca2≤600 μmol/L)时,ζ较大,无振荡现象 (图4)。从图4可见,随着CO2阶跃阶点的升高(Ca2>600 μmol/L),ζ急剧减小,振荡剧烈。Ca2对时间常数T的影响呈现单峰趋势关系,在Ca2=900 μmol/L时,T最大。另外,阶点对无阻尼自然振荡频率ωn和阻尼振荡频率ωd的影响不明显。
光强对CO2阶跃变化下茄子光合动态响应特征的影响
在处理Ⅲ的CO2阶跃变化 (15→1200 μmol/L)下,茄子Pn和Vc的动态进程也明显受光照水平的影响 (图5)。从图5可见,光合有效辐射较低 (PAR=300 μmol·quata/m2·s)时,茄子Pn和Vc无明显的振荡现象。当PAR=600 μmol·quata/m2·s时,Pn和Vc开始表现出振荡现象。随着PAR继续增加 (PAR从600至1800 μmol·quata/m2·s),Pn和Vc的振荡现象会愈加强烈。

图4 CO2阶跃阶点Ca2对茄子光合系统的阻尼系数ζ、无阻尼自然振荡频率ωn和阻尼振荡频率ωd(A),以及时间常数T(B)的影响Fig.4 Effect of CO2step order pointCa2on the damp coefficientζ,natural oscillation frequency for undamped presentωn,damped oscillation frequencyωd(A),and time constantT(B)of photosynthetic system in eggplant

图5 光强PAR对CO2阶跃变化(15→1200 μmol/L)下茄子光合速率Pn(A)和羧化速率Vc(B)动态进程的影响Fig.5 Effect of photosynthetically active radiationPARon the dynamic course of net photosynthesis rate Pn(A)and carboxylation rateVc(B)under the CO2step change from 15 to 1200 μmol/L in eggplant
将图5中Vc的动态进程进行振荡特征分析可见,光照水平对振荡特征的影响 (图6)与CO2阶跃阶点 Ca2的影响相似 (图 4)。从图 6 可见,PAR 较低 (PAR≤600 μmol·quata/m2·s)时,ζ较大,无振荡现象或有轻微的振荡。随着PAR升高 (PAR≥900 μmol/L),ζ急剧 减小,振荡也愈加剧烈。PAR对时间常数T的影响亦呈现单峰趋势关系,在PAR=600 μmol·quata/m2·s时,T最大。另外,PAR对无阻尼自然振荡频率ωn和阻尼振荡频率ωd的影响亦不明显。

图6 光强PAR对CO2阶跃变化(15→1200 μmol/L)下茄子光合系统的阻尼系数ζ、无阻尼自然振荡频率ωn和阻尼振荡频率ωd(A),以及时间常数T(B)的影响Fig.6 Effect of photosynthetically active radiation PAR on the damp coefficientζ, natural oscillation frequency for undamped presentωn,damped oscillation frequencyωd(A),and time constantT(B)of photosynthetic system under the CO2step change from 15 to 1200 μmol/L in eggplant
根据图1~图6中CO2阶跃起点Ca1、阶点Ca2、光强PAR对光合振荡的影响分析,将能够引起光合振荡的临界量Ca1o、Ca2o、PARo分别选 定 为 300 μmol/L、 900 μmol/L、500 μmol·quata/m2·s。据此计算的CO2阶跃扰动强度Dc与茄子光合振荡系统的阻尼系数ζ之间呈指数回归 关 系 , 即 ζ=0.7855e-0.0053Dc,其相关系数r=-0.858**,呈极显著负相关(P<0.01,图 7)。
讨 论
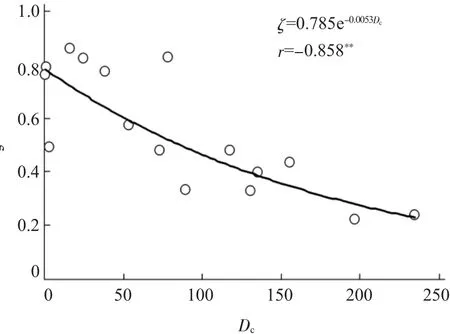
图7 茄子叶片光合系统的扰动强度Dc与阻尼系数ζ的关系Fig.7 Relationship between disturbed intensityDc and damp coefficientζof photosynthetic system in eggplant
光合的动态响应特征参数与动态光合的反馈控制机制
大气CO2浓度Ca骤然增加会引起羧化位点Cc的阶跃升高,进而引起Vc骤然增加。这会造成原稳态下底物池中ER(rubisco酶与RuBP复合体)的迅速消耗。而底物池中ER的补充速度一时较慢,会造成接续的Vc降低,从而表现出光合振荡现象。因此,CO2阶跃变化下光合振荡响应的机制与ER的原贮量消耗和再生补充不平衡有关。这在卡尔文环中相当于RuBP的消耗速率与再生速率不平衡问题[20],也就在卡尔文环中通过羧化产物PGA的还原继而再生RuBP过程表现为反馈控制机制[3,5]。
光合系统反馈控制动态生化模型是根据光合系统酶接触反应动力学参数构建的[15]。据此模型的CO2阶跃响应传递函数中的振荡单元,可以计算茄子动态光合的振荡特征参数,主要包括光合反馈系统的阻尼系数ζ和时间常数T(相当于无阻尼自然振荡频率ωn的倒数)。本研究发现,CO2阶跃变化对茄子光合系统的震荡特征参数ζ和T产生了明显的影响,而ωn和ωd受影响的程度则显著减小 (图2、4和6)。因此,动态光合的振荡发生及其振荡强度与光合反馈系统的ζ和T有关,亦即在卡尔文环中相当于RuBP的消耗速率与再生速率不平衡问题与光合反馈系统的ζ和T有关。据此推测,T应该与RuBP再生速率有关,ζ应该与RuBP底物池大小有关。但是,此推测有待研究探索。动态光合中影响CO2阶跃扰动的因子
在Farquhar、von Caemmerer和Berry[21,22]提出的C3植物光合作用稳态生化模型中,羧化速率Vc的限制因素有三[11]:1)在较低CO2浓度下,Vc受限于Rubisco的数量、动力学特性及其活性状态 (Wc);2)在中等CO2浓度下,Vc受限于与光系统电子传递有关的RuBP再生 (Wj);3)在高CO2浓度下,Vc受限于无机磷可利用量 (Wp)。可见,不同的CO2浓度和光强水平会使光合反馈系统处于不同的运转状态。但是,CO2从叶肉细胞扩散至叶绿体羧化部位的导度不同,影响Rd、Kc、Ko、Γ*和gi等的推算[23~25],对茄子动态光合过程的特征参数也将产生影响。因此推测,CO2阶跃变化引起的光合动态进程会依CO2阶跃梯度、阶跃起点、阶跃阶点的大小,以及光强水平的高低而影响到整个光合系统的状态。这在本研究中已经得到充分的证实。
同时,通过引入“CO2阶跃扰动强度”的概念探讨了CO2阶跃变化对茄子光合振荡程度的影响 (图7)。但是,这里初步选定的能够引起光合振荡的临界量 (Ca1o、Ca2o、PARo)可能与较大的CO2阶跃变化处理梯度有关,因为较小的CO2阶跃变化处理梯度需要光合仪双气路CO2稳衡调试、叶片光合稳衡调试,还需要CO2配气相配合,以及3次重复等,不仅增加了动态光合测定的工作量,而且也扩大了整个繁琐工作中每个环节存在的实验误差。另外,温度是直接影响植物光合系统酶促反应的重要因子之一,间接调控光合机构的运转状况。因此,CO2阶跃扰动强度的计算是否需要带入温度因子有待研究。
结 论
CO2阶跃变化下,光合系统阻尼系数ζ的减小和时间常数T的增加是引起茄子叶片光合速率Pn或羧化速率Vc发生振颤的主要原因,而无阻尼自然振荡频率ωn和阻尼振荡频率ωd的作用则不明显。
1. van Der Veen R.Induction phenomena in photosynthesis.Plant Physiol,1949,2:287~296
2.Rabinowitch EI.Photosynthesis and related processes(Vol.Ⅱ,part 2).New York:Interscience publishers,1956.537~1432
3.Laisk A,Oja V.Positive feedback and rhythmic phenomena in the pentosephosphate cycle of photosynthesis.Abstract of the Fourth International Biophysical Congress.Moscow.1972,sections XVI-XXV:62
4. Laisk A,Oja V.Photosynthesis at CO2and light saturation in limited by the reaction of rubilosediphosphate resynthesis.Proc Est Acad Sci,1976,25(2):146~150
5.Walker DA,Sivak MN,Prinsley RT,Cheesborough JK.Simultaneous measurement of oscillations in oxygen evolution and chlorophyll a fluorescence in leaf pieces.Plant Physiol,1983,73:542~549
6.Ogawa T.Simple oscillations in photosynthesis of higher planta.Biochim Biophys Acta,1982,68:103~109
7.高志奎,高荣孚,何俊萍,王 梅.环境因子对温室茄子光合非稳态性的影响.园艺学报,2005,32(4):624~627 Gao ZK,Gao RF,He JP,Wang M.Effect of environmental factors on unsteady-state photosynthesis in greenhouse Aubergine.Acta Horticul Sin,2005,32(4):624~627
8.Laisk A,Laarin P.Feedback control of the potential rate of photosynthesis.In:Margna U.Regulation of Plant Growth and Metabolism.Tallinn:Valgus,1983.135~150
9. Laisk A,Walker DA.Control of phosphate turnover as a rate-limiting factorand possible cause ofoscillations in photosynthesis:A mathematical model.Proc R Soc Lond B,1986,227:281~302
10.Laisk A,Walker DA.A mathematical model of electron transport. Thermodynamicnecessityforphotosystem Ⅱregulation:'light stomata'.Proc R Soc Lond B,1989,237:417~444
11.Laisk A,Eichelmann H.Towards understanding oscillations:A mathematical model of the biochemistry of photosynthesis. PhilTransR SocLond, 1989, 323:369~348
12.Mott KA,Jensen RG,O'leary JW,Berry JA.Photosynthesis and ribulose 1,5-bisphosphate concentrations in intact leaves ofXanthium strumariumL.Plant Physiol,1984,76:968~971
13.von Caemmerer S.Biochemical models of leaf photosynthesis.Australia:CSIRO publishing,2000.29~90
14.薛定宇,张晓华.控制系统计算机辅助设计——M A TLA B语言与应用(第2版).北京:清华大学出版社,2006.39~231 Xue DY,Zhang XH.Computer aided control systems design using matlab language(Second edition).Beijing:Tsinghua university press,2006.39~231
15.高志奎,薛占军,钱 稷,程 雪,何俊萍,高荣孚.光合作用的CO2阶跃响应动态生化模型.生物物理学报,2010,26(9):814~822 Gao ZK,Xue ZJ,Qian J,Cheng X,He JP,Gao RF.A dynamic biochemical model for photosynthetic response to CO2step changes.Acta Biophys Sin,2010,26(9):814~822
16.Kramer DM,Avenson TJ,Edwards GE.Dynamic flexibility in the light reactions of photosynthesis governed by both electron and proton transfer reactions.Trends Plant Sci,2004,9(7):349~357
17.Rascher U,Nedbal L.Dynamics of photosynthesis in fluctuating light.Curr Opin Plant Biol,2006,9:671~678
18.Nowak RS,Ellsworth DS,Smith SD.Functional responses of plants to elevated atmospheric CO2—— Do photosynthetic and productivity data from FACE experiments support early predictions? New Phytologist,2004,162:253~280
19.Toebert NE.Effeet of atmospherie CO2on Photosynthesis.In:Sybesrna C.Advances in photosynthesis researeh(Vol.IV).Boston:Laneaster,1984.81
20.Laisk A,Oja V.Dynamics of leaf photosynthesis:Rapidresponse measurements and their interpretations.Australia:CSIRO publishing,1998.58~88
21.Farquhar GD,von Caemmerer S,Berry JA.A biochemical model of photosynthetic CO2assimilation in leaves of C3species.Planta,1980,149(1):78~90
22.Farquhar GD,von Caemmerer S.Modeling of photosynthetic response to environmental conditions.In:Lange OL,Nobel PS,Osmond CB,Ziegler H.Physiological plant ecologyⅡ.Encyclopedia of plant physiology.New series,Vol.12B.Heidelberg:Springer-Verlag,1982.550~587
23.Ethier GJ,Livingston NJ.On the need to incorporate sensitivity to CO2transfer conductance into the Farquharvon Caemmerer-Berry leaf photosynthesis model.Plant Cell Environ,2004,27:137~153
24.Terashima I,Ono K.Effects of HgCl2on CO2dependence of leaf photosynthesis:Evidence indicating involvement of Aquaporinsin CO2diffusion across the plasma membrane.Plant Cell Physiol,2002,43:70~78
25.Yin X,Oijen V,Schapendonk HCM.Extension of a biochemicalmodelforthe generalized stoichiometry of electron transportlimited C3photosynthesis. PlantCell Environ,2004,27:1211~1222
Effect of CO2Step Changes on Characteristic Parameters of Dynamic Photosynthesis in Eggplant
XUE Zhanjun1,GAO Zhikui1,WANG Mei1,LV Guiyun1,DU Shuang1,GAO Rongfu2
1.College of Horticulture,Agricultural University of Hebei,Baoding 071001,China;
2.College of Biological Sciences and Technology,Beijing Forestry University,Beijing 100083,China
This work was supported by grants from The National Natural Science Foundation of China(30571267);Science and Technology Research Projects of China(2004BA525B14)
Oct 18,2010 Accepted:Dec 28,2010
GAO Zhikui,Tel:+86(312)7528312,E-mail:gaozhikui2005@163.com
With the constitution of dynamic biochemistry model containing a key of feedback controlled system,which was on the basis of oscillation unit in the transfer function,the characteristic parameters of oscillation in dynamic photosynthesis by CO2step-transformation were analyzed in solar greenhouse eggplant treated by combination of CO2step starting point(Ca1)and order point(Ca2)with different photosynthetically active radiation(PAR).The results showed that dynamic course of oscillation of photosynthetic rate(Pn)and carboxylation velocity(Vc)which was translated from Pnthrough stable photosynthesisparameterswas significantly violent due to the increase of time constant(T)and decrease of damp coefficient (ζ),natural oscillation frequency for undamped present(ωn)and damped oscillation frequency(ωd)of photosynthetic system.Analogously,under the condition ofCa2≥900 μmol/L andPAR≥900 μmo·lquata/m·2s,the phenomena of oscillation was more marked accompanied the lowerζ.Consequently,the photosynthetic oscillation could occur in combination of lowerζand upperT.Conversely,it could not display in combination of upperζ and lowerT.However,the moderateζand high(or higher)Twere the critical conditions needed for occurring of photosynthetic oscillation.In our experiment,the critical quantity of photosynthetic oscillation for Ca1,Ca2andPARwas 300 μmol/L,900 μmol/L and 500 μmol·quata/m2·s,respectively.Meanwhile,the negative relationship(r=-0.858**)between disturbed intensity(Dc)by CO2step-transformation and damp coefficient(ζ)of photosynthetic oscillation system can be described by exponential equation.
Dynamic photosynthesis;Oscillation;CO2step change;Damp coefficient;Oscillation frequency;Time constant
2010-10-18;接受日期:2010-12-28
国家自然科学基金项目 (30571267),国家科技攻关计划项目 (2004BA525B14)
高志奎,电话:(0312)7528312,E-mail:gaozhikui2005@163.com
Q945.11,S641.1
猜你喜欢
黑龙江大学自然科学学报(2021年4期)2021-11-19 07:05:06
重型机械(2019年3期)2019-08-27 00:58:52
天津教育·下(2018年9期)2018-07-13 08:25:47
中国有色金属学报(2018年6期)2018-07-09 08:55:42
化工技术与开发(2015年6期)2015-01-29 02:11:46
物探化探计算技术(2014年1期)2014-06-27 01:32:50
华东理工大学学报(自然科学版)(2014年2期)2014-02-27 13:48:40
电力自动化设备(2013年11期)2013-09-18 02:55:14
电气电子教学学报(2013年1期)2013-07-05 09:41:14
电气电子教学学报(2012年1期)2012-10-12 03:27:48