
红尾伯劳适应性巢址选择研究
2011-07-09 01:37徐纯柱郭自荣
东北农业大学学报 2011年8期
徐纯柱,郭自荣
(东北农业大学生命科学学院,哈尔滨 150030)
随着城市化进程的不断推进,城市中的高大建筑和硬化路面逐渐取代了自然绿地景观[1]。城市生态系统所呈现的生境破碎化现象严重影响了城市野生鸟类的种群数量及分布,但有些野生鸟类也在人类活动中表现出了一些适应性特征。因此,探讨城市化程度与城市野生鸟类之间的共存机制对于维护城市生态系统的平衡具有重要意义[2]。
鸟类巢址选择研究是鸟类学研究中的一个重要分支,在很多濒危鸟类中都有很好的研究成果[3-5]。是分析鸟巢及其周边的生态因子在鸟类选择巢址过程中的作用和地位,从而揭示鸟类选择该处筑巢的原因和主导因素。鸟类巢址选择的质量高低与其繁殖成功率有直接关系[6-8]。因此,研究野外和城市环境中鸟类巢址选择的差异性,对于探讨野鸟对异质性环境的适应能力,评价城市生态建设以及鸟类资源的保护、管理和利用等方面均具有重要意义。
红尾伯劳(Lanius cristatus)是一种被列入《中日保护候鸟及其栖息环境的协定》的农林益鸟。国内分布有4个亚种:普通亚种(L.c.lucionensis)、指名亚种(L.c.cristatus)、东北亚种(L.c.confusus)及日本亚种(L.c.superciliosus),黑龙江省境内仅有东北亚种繁殖[9]。有关红尾伯劳的生态学研究成果较多[10-12],但大多针对于繁殖习性方面的报道,对于其在野外和城市环境中巢址选择的对比研究尚未见详细报道。鉴于此,笔者于2007~2010年对黑龙江省帽儿山和横道河子两处实习基地以及哈尔滨市内东北农业大学校园内分布的红尾伯劳巢址进行了研究。
1 研究区域与方法
1.1 研究地概况
横道河子和帽儿山都是东北农业大学动物学野外实习基地,地理坐标分别为 129°02′08″E,44°48′47″N 和 127°30′~127°34′E,45°20′~45°25′N。分属于长白山系支脉张广才岭东麓和西北坡余脉。
两地区的植被均属于长白山植物区系,具有东北东部地区典型的温带天然次生林。林冠下丰富的灌木植被和充足的天然水源,为鸟类栖息、繁殖提供了良好的生境条件。
东北农业大学位于哈尔滨市内,校区整体植被覆盖面积均超过50%,且植被组成中乔木以旱柳(Salix matsudana)、春榆(Ulmus japonica)、大果榆(U.macrocarpa)、白桦(Betula platyphylla)、银白杨(Populus alba)、糖槭(Acer negundo)、红皮云杉(Picea koraiensis)和暴马丁香(Syringa reticulata)等为主;灌木主要以日本小檗(Berberis thunbergii)、红瑞木(Cornus alba)和紫丁香(Syringa oblata)等为主;草本以禾本科(Poaceae)和菊科(Asteraceae)植物为主。
1.2 方法
1.2.1 数据收集方法
统计巢址来源包括:2007~2010年6~7月在帽儿山和横道河子两处动物学实习基地发现红尾伯劳巢24个,东北农业大学校园内发现红尾伯劳巢25个。以巢址为中心做2 m×2 m的样方生境因子调查,具体参数包括:营巢树种,巢向,巢树高度,胸径,巢枝直径,巢位高度,植被盖度,距房屋最近距离(m),距道路最近距离(m),距水源最近距离(m),距垃圾桶最近距离(m),隐蔽度,人为活动等。
其中隐蔽度分8个等级:在巢周围8个方向观察鸟巢,8个方向均无法见到,定义为8级;7个方向均无法见到,定义为7级;以此类推分成1~8级。
人为活动分5个等级:距巢树5 m范围内平均每分钟通过的人数>40人,定义为5级;距巢树5 m范围内平均每分钟通过的人数<30人,定义为4级;距巢树5 m范围内平均每分钟通过的人数<20人,定义为3级;距巢树5 m范围内平均每分钟通过的人数<10人,定义为2级;距巢树5 m范围内平均每分钟通过的人数<1人,定义为1级;巢向指巢在营巢树上的方向。
1.2.2 数据分析
野外数据及资料经过初步整理后,对样方内各植被的名称等文字型因子及9项数值型因子运用频次分析法进行粗略统计。采用因子分析(Factor analysis)的主成分法(Principal components)对相关变量进行主成分分析,筛选出影响红尾伯劳巢址选择的主要生境因子。然后对野外和城市环境中红尾伯劳巢的主要生境因子进行均值比较及Q型聚类分析。所有数据利用SPSS 17.0统计分析软件进行处理。全文数据用平均值±标准误表示。
2 结果与分析
2.1 校园环境红尾伯劳巢址选择主要因子筛选
对东北农业大学校园内红尾伯劳巢址数值型参数主成分分析(见表1)。结果表明,前3个主成分特征值均大于1,累积贡献率达72.26%,说明前3个主成分基本包含了10个参数的总信息量。提取前3个主成分并计算各变量特征向量(见表2)。从表2可知,第一主成分的巢位高度、巢树高度、人为活动和距道路最近距离因子载荷较大,但由于巢位高度与巢树高度(r=0.899,P<0.01)、人为活动(r=0.546,P<0.01)、距道路最近距离(r=-0.563,P<0.01)呈现强正或负相关,因此提取第一主成分的巢位高度因子。提取第二主成分中载荷较大的隐蔽度因子和第三主成分中的距房屋最近距离和植被盖度因子。
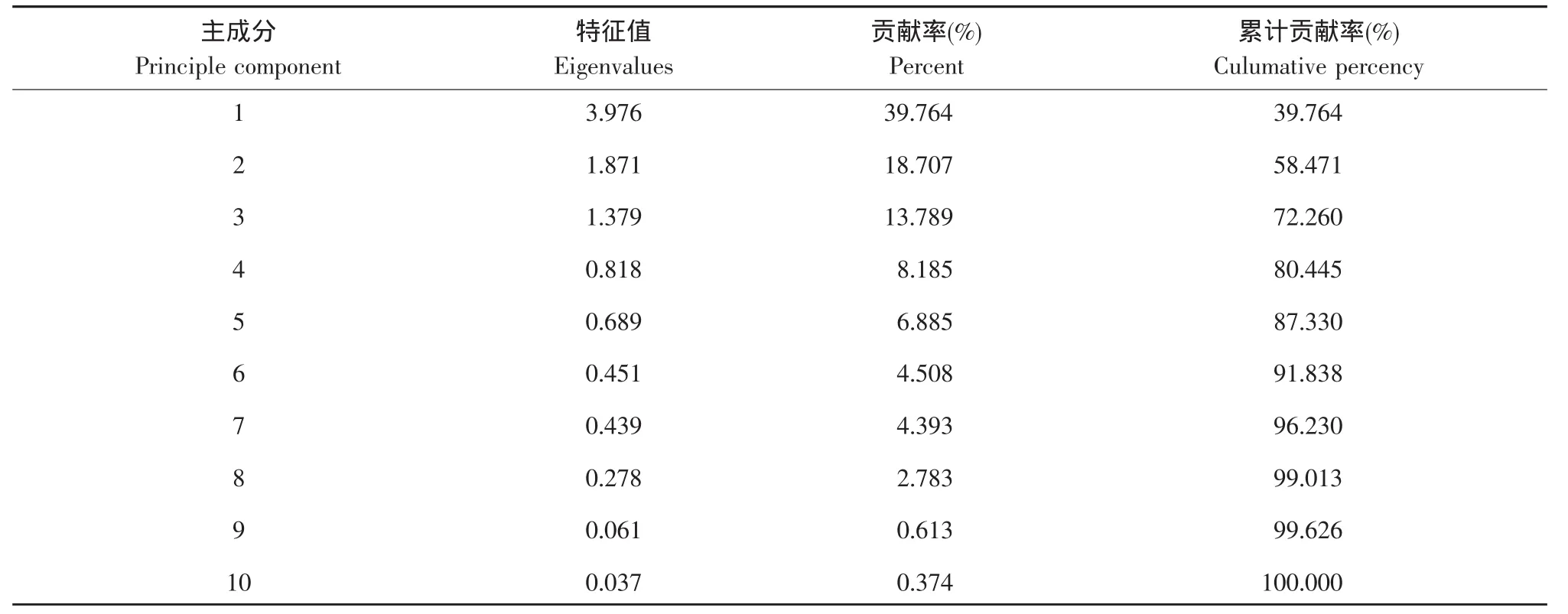
表1 校园内红尾伯劳巢址选择各主成分特征值Table 1 Eigenvalues of principle components for nest-site selection of the red-tailed shrike in campus environment
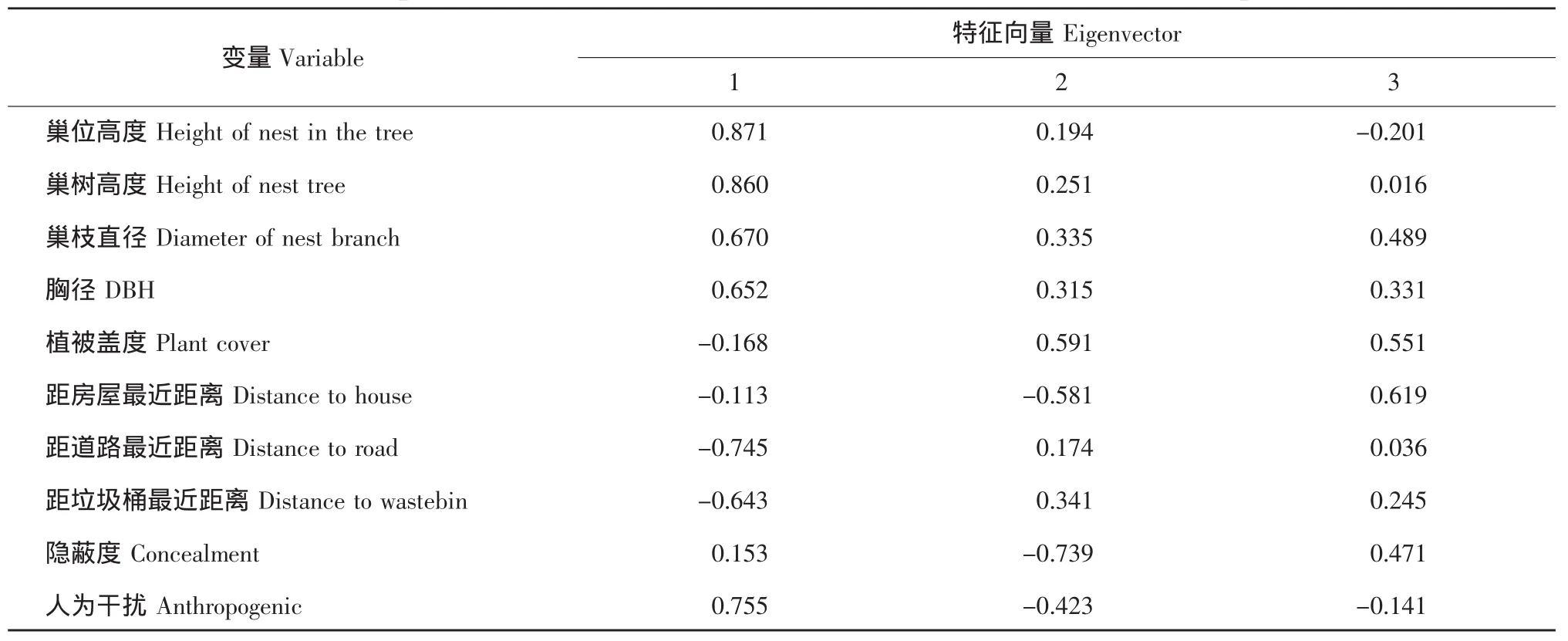
表2 校园红尾伯劳巢址选择的转置因子成分矩阵Table 2 Rotated component matrix for nest-site selection of the red-tailed shrike in campus environment
2.2 野外环境红尾伯劳巢址选择主要因子筛选
对两实习基地红尾伯劳巢址数值型参数主成分分析(见表3)。结果表明,前3个主成分特征值均大于1,累积贡献率达81.91%,说明前3个主成分基本包含了8个参数的总信息量。提取前3个主成分并计算各变量特征向量(见表4)。即第一主成分的巢位高度、巢树高度、隐蔽度、距道路最近距离、胸径和巢枝直径因子载荷较大,但由于巢位高度与巢树高度(r=0.718,P<0.01)、巢枝直径(r=0.742,P<0.01)、胸径(r=0.615,P<0.01)呈现强正相关,且前者决定后者,因此提取第一主成分的巢位高度、隐蔽度和距道路最近距离等因子。第二主成分中,距水源最近距离因子载荷较大,将其提取。提取第三主成分的植被盖度因子。
2.3 校园与野外环境巢址样方对比结果
红尾伯劳校园巢址样方(n=25)与野外巢址样方(n=24)3个主要生态因子测量与比较结果见表5。结果显示:两种环境中的巢址选择在巢位高度、植被盖度、隐蔽度3个变量间均存在极显著差异(P<0.01)。红尾伯劳在校园环境中选择巢址的巢位高度和隐蔽度均较野外环境中的巢址高,而植被盖度明显低于野外环境中的红尾伯劳巢址。
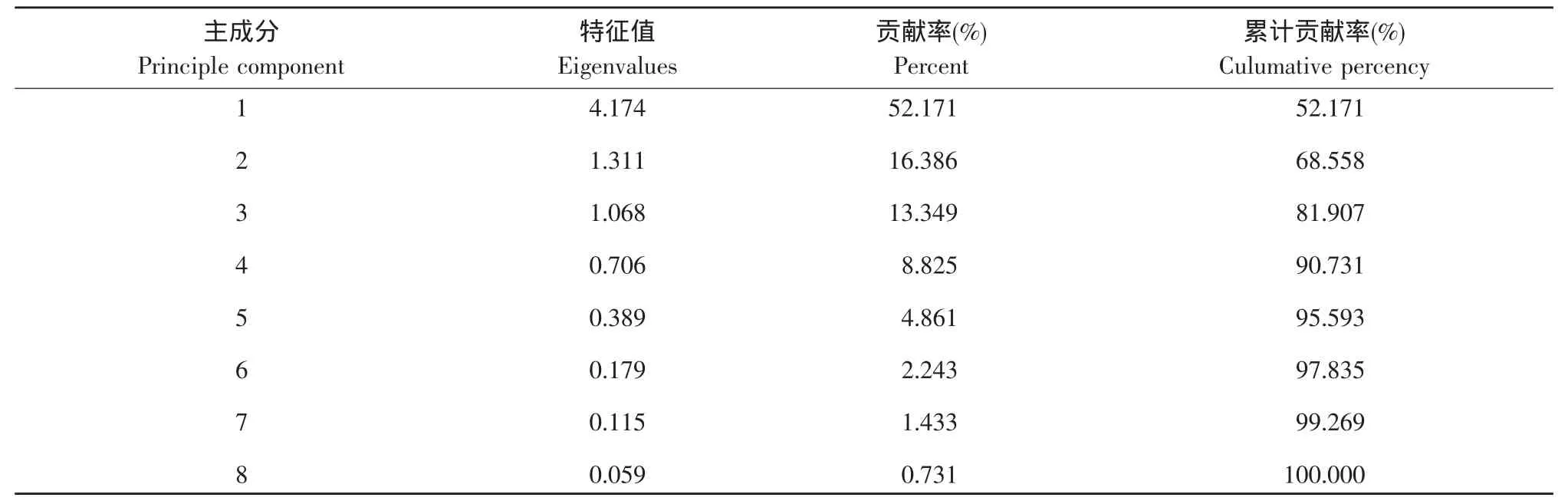
表3 野外红尾伯劳巢址选择各主成分特征值Table 3 Eigenvalues of principle components for nest-site selection of the red-tailed shrike in field environment
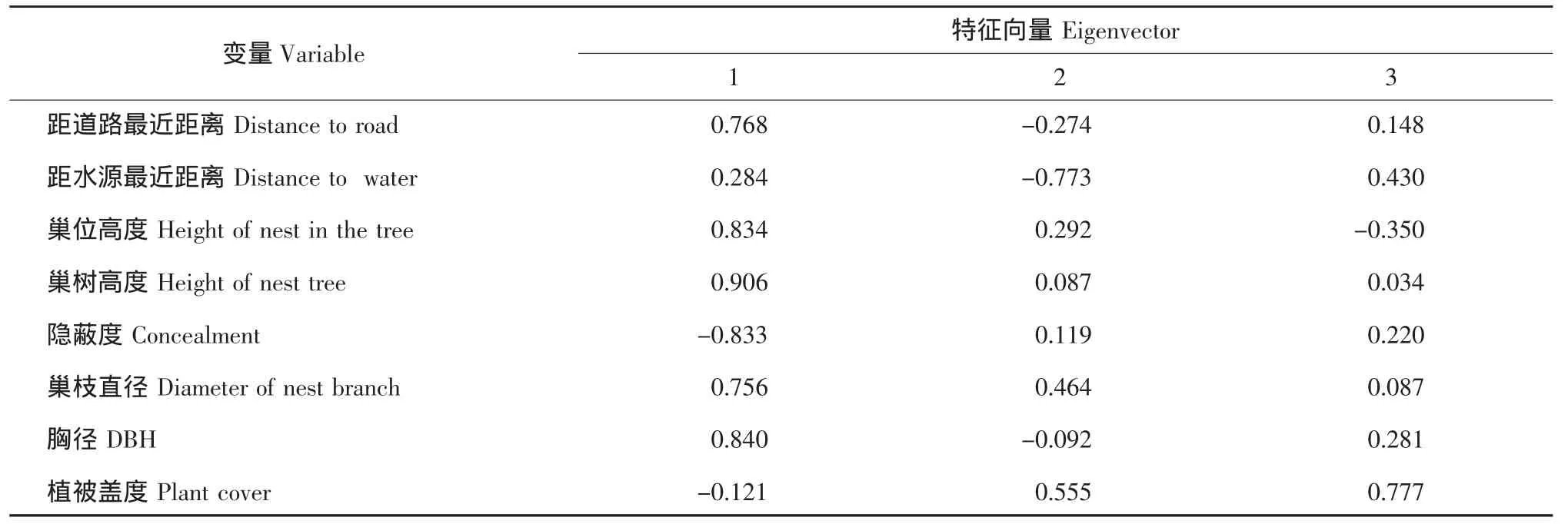
表4 野外红尾伯劳巢址选择的转置因子成分矩阵Table 4 Rotated component matrix for nest-site selection of the red-tailed shrike in field environment
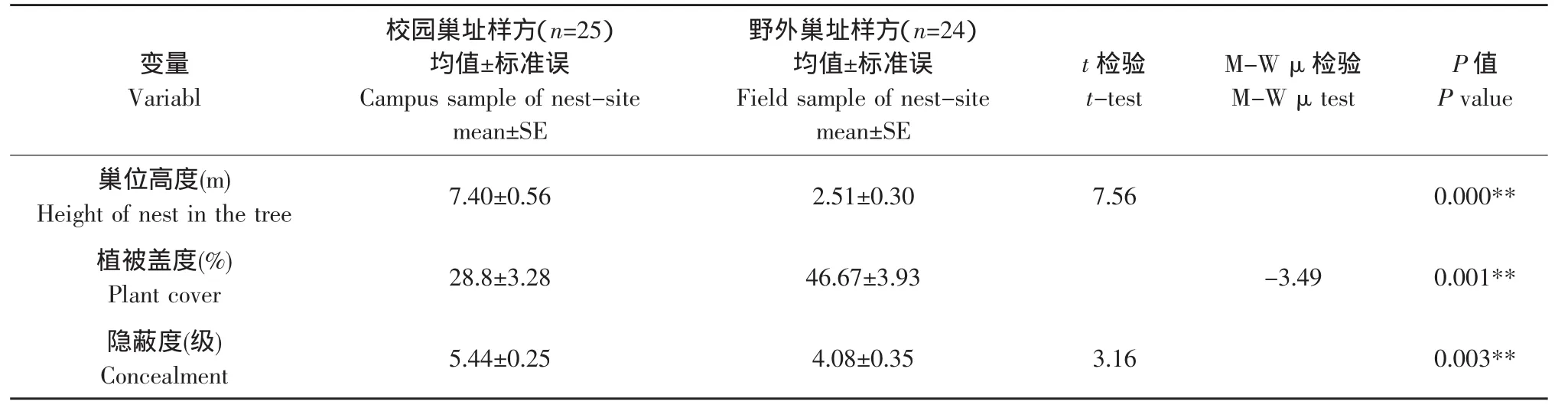
表5 红尾伯劳校园与野外巢址样方特征比较Table 5 Characteristic comparisons between the campus and field samples of red-tailed shrike
同时,对全部红尾伯劳巢址进行的Q型聚类结果显示:49个巢址大致可清晰地分为两大类,分属校园环境和野外环境巢址选择类型,其中16、18、22、47号巢例外(见图1)。
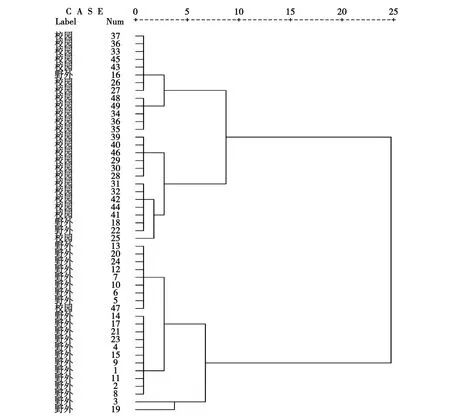
图1 校园与野外红尾伯劳巢址Q型聚类分析Fig.1 Analysis of Q cluster on red-tailed shrike nest-site between the campus and field environment
3 讨论与结论
巢址选择对于鸟类生存和繁殖具有重要意义,巢址质量的优劣将直接影响当年的繁殖成功率,从而间接影响种群动态和群落组成[13]。因此,很多鸟类在巢位选择过程中都倾向于选择那些能使其繁殖成效最大而存活代价最小的营巢生境[14]。本研究结果表明,无论校园还是野外环境,巢位高度因子均为影响红尾伯劳巢址选择的最主要因素,其贡献率分别达39.76%和52.17%,这与很多城市鸟类[15],特别是四川南充市内校园环境下红尾伯劳巢址选择情况一致[10]。其次是干扰因子(校园种群)和营巢树胸径(野外种群),最后均是隐蔽度因子,这与大多数鸟类都不尽相同。原因可能在于校园环境受人类活动影响较强,红尾伯劳必须首先选择距离道路较远、人为活动较少的区域营巢,其次再考虑隐蔽度因子,因为在干扰强度大的位置筑巢,要求隐蔽度高,从而导致光照因子及行动等方面均不利于亲代的成功繁殖。而野外种群在选择隐蔽度因子前首先选择营巢树胸径因子,推测巢的稳固性占主导地位,野外风力较校园大很多,因此野外环境的红尾伯劳必须选择胸径较大、巢位较低的位置营巢,以确保巢址稳定性,加之野外环境中的人为干扰程度较低,隐蔽度相对较差,这也是在野外环境较校园内容易发现红尾伯劳巢的一个重要原因。
此外,通过对红尾伯劳巢址Q型聚类分析发现,49个巢址大致分成两大类,分属校园环境和野外环境巢址选择类型,呈现显著的趋异化,即野外环境趋于较低巢位高度和隐蔽度以及较高植被盖度,而校园环境则趋于较高巢位和隐蔽度以及较低植被盖度。推测校园环境受人类活动影响较强,风力较小。红尾伯劳将巢位高度上移,适当降低隐蔽度是鸟类有效适应城市环境变化的一种方式。而野外环境受人类活动影响相对较小,但自然风力较大。红尾伯劳将巢位高度下移,确保巢的稳固性并适当增加巢隐蔽度是鸟类适应野外环境成功繁殖的一种策略。由此可见,红尾伯劳在校园和野外环境中的巢址选择采取了不同模式的隐蔽条件组合,在两种环境中巢址选择的分异不仅表现在单个生境因子的数值差异,更表现为群体隐蔽对策的不同。
[1]Fernández-Juricic E,Jokim ki J.A habitat island approach to conserving birds in urban landscapes:case studies from southern andnortherneurope[J].Biodiversityand Conservation,2001,10(12):2023-2043.
[2]Niemel J.Ecology and urban planning[J].Biodiversity and Conservation,1999,8(1):119-131.
[3]闫永峰,刘迺发.盐池湾自然保护区喜马拉雅雪鸡的巢址选择[J].干旱区地理,2011,34(1):160-164.
[4]李宏群,廉振民.陕西黄龙山自然保护区褐马鸡营巢生境[J].林业科学,2010,46(10):186-190.
[5]田军东,董瑞静,路纪琪.郑州大学新校区喜鹊巢址选择研究[J].河南师范大学学报:自然科学版,2009,37(5):116-118.
[6]闫永峰,刘迺发.东大山自然保护区喜马拉雅雪鸡(Tetraogallus himalayensis)的巢址选择[J].生态学报,2009,29(8):4278-4284.
[7]Robertson G Y.Factors affecting nest site selection and nesting success in the common eider(Somateria mollissima)[J].Ibis,1995,137(1):109-115.
[8]刘学昌,吴庆明,邹红菲,等.丹顶鹤(Grus japonensis)东,西种群巢址选择的分异[J].生态学报,2009,29(8):4483-4491.
[9]陈服官,罗时有,郑光美,等.中国动物志·鸟纲·第九卷·雀形目(太平鸟科-岩鹨科)[M].北京:科学出版社,1998:23-28.
[10]青云,周友兵,杨容,等.红尾伯劳的巢址选择与营巢行为[J].动物学杂志,2004,39(4):77-80.
[11]张绳祖,王定乾,都伟,等.红尾伯劳的繁殖习性[J].西北师范大学学报:自然科学版,1986(2):57-63.
[12]郑光美,魏潮生.红尾伯劳的繁殖习性[J].动物学报,1973,19(2):182-189.
[13]张微微,马建章,李金波.骨顶鸡(Fulica atra)的巢址选择研究[J].东北师大学报:自然科学版,2010,42(3):114-120.
[14]Valkama J,Korpimaki E,Utilization T P.Diet and reproductive success in the kestrel in a temporally and spatially heterogeneous environment[J].Ornis Fennica,1995,72:49-61.
[15]陈侠斌,何静,张薇.北京高校喜鹊巢址选择的主要生态因素[J].四川动物,2006,25(4):855-857,861.
猜你喜欢
天津教育(2023年2期)2023-03-14
学与玩(2022年9期)2022-10-31
廉政瞭望·下半月(2022年4期)2022-05-12
浙江林业(2022年3期)2022-04-12
红领巾·探索(2021年9期)2021-09-14
文苑(2020年12期)2020-04-13
小太阳画报(2019年1期)2019-06-11
课外生活(小学1-3年级)(2019年4期)2019-05-04
现代园艺(2017年13期)2018-01-19
电子制作(2017年14期)2017-12-18
